王建生 张杏锋 张学洪 吴炽珊 冯健飞


摘要:以李氏禾(Leersia hexandra Swartz)为例,研究了腐殖酸(humic acid,HA)和铬(Cr)相互作用对李氏禾生长及光合生理的影响。结果表明:在同一Cr浓度处理下,李氏禾株高、地上部干质量、叶绿素a含量、叶绿素b含量、总叶绿素含量随HA浓度的增大呈先增大后减小的趋势。在同一腐殖酸浓度处理下,Cr100、Cr200的株高、地上部干质量、地下部干质量、叶绿素a含量、叶绿素b含量、初始荧光F0、大荧光Fm较Cr0呈增大的趋势。初始荧光F0、最大荧光Fm、表观电子传递速率ETR在HA0和HA5处理下随Cr浓度的增加明显增加。而最大光化学效率F0/Fm、光化学淬灭系数(qp)、非光化学淬灭系数(qN)在HA5、HA10处理下,随Cr浓度的增加有所下降。可见,适量Cr和腐殖酸能够刺激植物生长,李氏禾在逆境胁迫下具有较强的适应性和耐受性。
关键词:李氏禾;铬污染;腐殖酸;光合生理特征;适应性;耐受性
中图分类号: X53;X173? 文献标志码: A? 文章编号:1002-1302(2019)05-0263-05
收稿日期:2017-10-21
基金项目:广西科学研究与技术开发项目(编号:桂科转1599001-1);广西特聘专家专项;广西危险废物处置产业化人才小高地项目。
作者简介:王建生(1990—),男,山东汶上人,硕士研究生,主要从事重金属污染修复。E-mail:wjs_0506@163.com。
通信作者:张杏锋,博士,副教授,主要从事重金属污染治理研究工作。E-mail:zhangxf@glut.edu.cn。
随着对采矿、制革、冶炼、电镀、污水灌溉等行业的大力发展,进而引起了铅、汞、镉、铬等一大批重金属污染物进入大气、水和土壤,造成了严重的环境污染问题[1]。而土地作为不可再生资源,是人类生活生存和自然环境发展的基础,所以土壤重金属污染问题普遍引起人们的关注[2]。铬(Cr)污染不仅会导致土壤生产能力的下降,而且还可以通过根部的吸收、迁移转化到农作物根茎叶及果实中去,通过食物链进入到人体内,进而危害人的身体健康[3]。因此,如何有效地将土壤中重金属去除又不污染其他资源是研究重点。植物修复技术被认为是非常理想的污染土壤原位治理技术[4-5]。李氏禾(Leersia hexandra Swartz)属多年生禾本科(Gramineae)草本植物,作为国内首次被发现的Cr超富集植物,在土壤Cr污染修复和治理上具有重要的利用价值[6]。
腐殖酸(humic acid,HA)是一种主要由动植物残体通过各种生物和非生物的降解、缩合等作用形成的一种天然有机高分子聚合物[7-8]。腐殖酸通过与重金属发生络(螯)合、吸附和氧化还原等反应,改变土壤形态和土壤对重金属的吸持力,影响有效性和植物的吸收[4]。腐殖酸对植物生长具有积极作用,可以促进植物营养物质吸收,调节植物的生理代谢,提高酶活性,同时还可以增加土壤微生物的总量。闫双堆等研究表明,在土壤中施腐殖酸类物质可以提高土壤中有机结合态汞的含量,使土壤中汞的挥发量有效降低[9]。腐殖酸可以促进大豆可交换态和碳酸盐形式转移到有机形式,表明大分子腐殖酸可以促进重金属从易分解转化为生物可利用部分[10]。在香根草中应用腐殖酸,提高了植物的铜摄取量及其在根中的积累量。研究表明,适量的腐殖酸能够促进植物种子的生长,使种子发芽率提高、萌发时间缩短、幼苗长势增加;腐殖酸类物质对植物根系的发育、提高和保持具有明显效果[11]。腐殖酸对湿生的重金属超富集植物的影响未见报道。本研究以李氏禾为例,研究了在Cr和腐殖酸下,李氏禾生长和生理生化指标的变化规律,进一步探究李氏禾耐受性[12]。
1 材料与方法
1.1 供试材料
1.1.1 供试植物和土壤供试 李氏禾于2016年6月中旬取自桂林市雁山区水田。盆栽试验用土2016年5月中旬采自广西桂林市雁山区未受污染的农田。
1.1.2 植物培养与处理 本试验于2016年6月20日在桂林理工大学内进行,采用2因素设计进行试验,其中1个因素为Cr的浓度,设置2个Cr浓度污染水平,每盆装晾干磨碎土2.5 kg,按含0、100、200 mg/kg(分别用Cr0、Cr100和Cr200表示)、重铬酸钾(K2Cr2O7)分别加入混匀,重复3次;另1个因素为腐殖酸浓度,腐殖酸采用3个处理、1个对照,按含0、5、10、20 mg/kg腐殖酸(分别用HA0、HA5、HA10和HA20表示)加入盆中混匀,3组平行。然后每盆中移栽8~10株李氏禾。持续处理3个月时间,定期浇水,每次浇水处理一样,保持土壤湿润,试验过程中除了重金属浓度和腐殖酸浓度不同以外,其他条件保持一致。
1.2 测定方法和数据分析
1.2.1 叶绿素含量测定 培养90 d后,从每盆中选取1株植株的顶部第3张或第4张叶片用于测定叶绿素含量,采用95%乙醇提取法测定叶绿素含量[13],用分光光度计在波长665、649、470 nm下分别测定吸光度,然后按公式计算各叶绿素含量。
1.2.2 叶绿素荧光参数测定 采用便携式调制叶绿素荧光仪(PAM-2500)于20:00—24:00時测定自然条件下李氏禾的叶绿素荧光参数。每盆选取1株长势一致的健康植株进行测量,从植物顶端数第4张至第5张叶片中部进行测量,每隔20 s照射饱和脉冲光。测量指标包括光下最大荧光(Fm′)、光下最小荧光(F0′)、最小荧光(F0)、最大荧光(Fm)、表观光合电子传递速率(ETR)、最大光化学效率(Fv/Fm)、光化学淬灭系数(qp)和非光化学淬灭系数(qN),所有参数均由仪器自动测得,依次计算出PSⅡ潜在活性(Fv/F0)。
1.2.3 生长指标 试验结束后,采用直尺测量植株高度,然后收获植物后用天平测定。将收获的植物样品分为地上、地下2个部分,分别用自来水充分冲洗,去除表面上泥土和污物,然后用去离子水冲洗,放入恒温箱在105 ℃下杀青 30 min,最后置于70 ℃的烘箱中烘干至恒质量,测定各部分物质的含量。
1.2.4 数据处理 所得数据应用SPSS 20.0统计分析软件分析各处理间的差异显著性,平均值间的比较采用单因素方差分析方法,显著性水平设定为a=0.05。
2 结果与分析
2.1 重金属和腐殖酸对李氏禾生长的影响
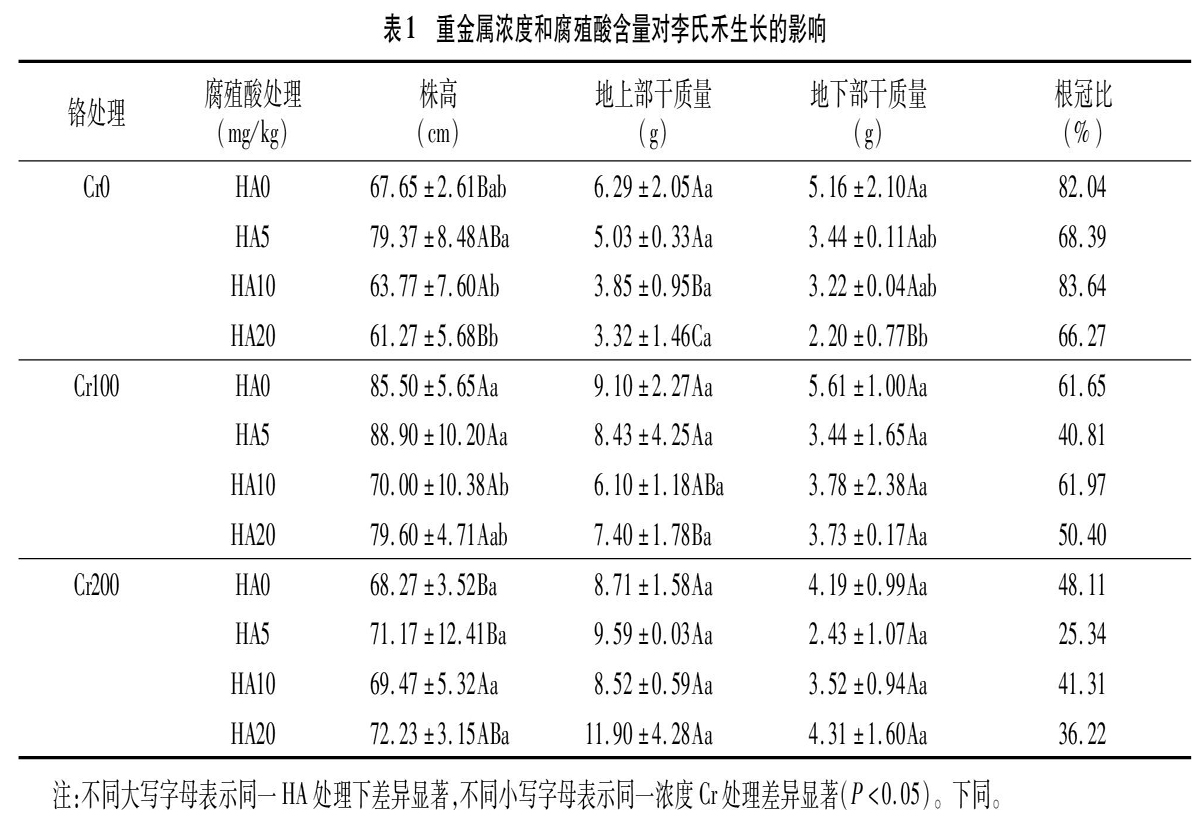
不同处理下植物株高如表1所示,在同一浓度HA0和HA20处理下,植物株高在Cr100和Cr200处理下明显高于Cr0处理;但是在同一浓度HA5和HA10处理下,植物株高在Cr100和Cr200处理下与Cr0处理没有显著差异。在同一浓度Cr100处理下,李氏禾株高在HA10和HA20处理明显低于HA0处理。可以看出,适量的腐殖酸在清洁土壤和Cr污染土壤下可以促进植物株高的生长。
植物地上部干质量是反映环境因素对植物生长的重要指标。从表1可以看出,在同一浓度腐殖酸处理时,植物地上部干质量随着Cr浓度的增加而增加。在同一Cr0处理下,植物地上部干质量在HA5、HA10和HA20处理下分别比HA0处理下降20.03%、38.79%和47.22%。但是,在同一浓度Cr200处理下,HA5和HA20处理下腐殖酸的加入使植物地上部干质量分别比HA0增加10.10%和36.62%。可以看出,腐殖酸在清洁土壤和低浓度Cr污染时会抑制植物生长,但在高浓度Cr污染时会刺激植物生长。以上结果说明腐殖酸与Cr对植物生长具有交互作用,随着土壤浓度不同,其交互作用不同,在低浓度时表现出拮抗作用,在高浓度时表现出协同作用。
植物地下部干质量反映植物根系生长情况。从表1可以看出,在同一浓度HA0处理下,植物地下部干质量随着Cr浓度的增加先增加后减少;而同一浓度HA10处理下,植物地下部干质量随Cr浓度的增加变化不明显;而在同一浓度HA20处理下,植物地下部干质量在Cr100和Cr200处理下显著高于Cr0,分别比Cr0增加69.55%和95.91%。在同一浓度Cr0处理下,腐殖酸的加入明显降低植物地下部干质量,HA5、HA10、HA20分别比HA0下降33.33%、37.60%、57.36%。而在同一浓度Cr0、Cr100、Cr200处理时,除Cr200的HA20处理外,植物地下部干质量在HA5、HA10、HA20下较HA0降低。以上结果说明,高浓度Cr含量会降低植物地下部根系的生长,而腐殖酸在清洁土壤上抑制植物根系生长。
根冠比反映植物地下部分与地上部分的相关性,高则根系机能活性强,低则弱。从表1可以看出,同一浓度腐殖酸处理下,植物根冠比随着Cr浓度的增加而减少;而同一浓度HA5处理下明显下降,植物根冠比Cr100和Cr200处理下分别比Cr0降低58.30%和66.12%;在同一浓度Cr0处理下,植物根冠比随腐殖酸浓度的增加先增大后减少;而在同一浓度Cr100和Cr200处理下,植物根冠比在HA5、HA10、HA20处理下较HA0有所下降。可以看出,适量的腐殖酸在清洁土壤中可以促进植物根冠比。腐殖酸在Cr污染土壤下却抑制根冠比,腐殖酸和Cr表现为协同作用。
2.2 重金属和腐殖酸对李氏禾叶绿素的影响
叶绿素是植物光合作用的主要色素,其含量的高低一定程度上反映植物光合作用能力的强弱,叶片中叶绿素含量低,其光合作用则弱,对植物的生长不利[14]。某些金属离子在一定质量分数范围内会对叶绿素的合成具有促进作用。
从表2可以看出,在同一浓度HA0处理下,植物叶绿素a含量在Cr100和Cr200处理下明显高于Cr0处理,分别比Cr0处理增加9.29%和19.47%;在同一濃度HA5、HA20处理下,植物叶绿素a含量在Cr100和Cr200处理下较Cr0有所增加但不显著。在Cr0处理下,腐殖酸的加入促进了植物叶绿素a的增长,植物叶绿素a含量在HA5、HA10、HA20处理分别比HA0处理增加16.37%、27.88%、19.91%;在Cr100处理下,植物叶绿素a含量在HA5、HA10、HA20处理下分别比HA0处理增加 23.48%、10.93%、12.15%,其中在HA5下达到峰值;在Cr200处理下,植物叶绿素a含量在HA5、HA10、HA20处理下分别比HA0处理下增加8.89%、9.26%、8.15%。可以看出腐殖酸无论是在清洁土壤还是Cr污染土壤都能够促进植物叶绿素a含量的增加。说明适量的Cr和腐殖酸对叶绿素含量增加具有一定的促进作用。
从表2可以看出,在同一浓度腐殖酸处理下,植物叶绿素b含量随着Cr浓度的增加而增加;在HA0处理下,植物叶绿素b含量在Cr100和Cr200处理下分别比Cr0处理增加20.69%、152.87%。在Cr0和Cr100处理下,腐殖酸的加入明显增加植物叶绿素b含量;在Cr0处理下,植物叶绿素b含量在HA5、HA10、HA20处理下分别比HA0增加28.74%、63.22%、44.83%;而在Cr200处理下,HA5、HA10处理下腐殖酸的加入明显增加了植物叶绿素b含量,植物叶绿素b含量在HA5、HA10、HA20处理下分别比HA0处理增加5273%、43.64%、2.73%,HA5处理下叶绿素b含量达到最大值。由此可以看出,腐殖酸在清洁土壤和Cr污染土壤下都能刺激植物叶绿素b含量增加。以上结果说明腐殖酸与Cr对李氏禾叶绿素b含量增加具有协同作用。
叶绿素a/b值反映了植物对光能的利用程度。从表2中可以看出,同一浓度腐殖酸处理时,植物叶绿素a/b随着Cr浓度的增加而降低;而在HA5和HA10处理下,植物叶绿素a/b值随Cr浓度的增加明显下降;HA5处理下,植物叶绿素a/b值在Cr100和Cr200处理下分别比Cr0处理下降29.66%和61.02%。在同一浓度Cr处理下,除Cr200 HA20处理,植物叶绿素a/b值随腐殖酸浓度的增加而下降;而在Cr200处理时,HA5、HA10处理植物叶绿素a/b值较HA0处理明显下降,在HA5、HA10和HA20处理下分别比HA0处理下降51.58%、48.42%和23.68%。由此可以看出,腐殖酸在清洁土壤和Cr污染时抑制植物叶绿素a/b值,说明腐殖酸和Cr对植物叶绿素a/b值表现为协同作用。叶绿素a/b值的降低,说明其捕获、转化光能的能力下降,同时也可以看出叶绿素a受破坏的程度较叶绿素b严重。
从表2中可以看出,在同一浓度腐殖酸处理下,植物胡萝卜素含量随着Cr浓度的增加而降低。在HA0处理下,植物胡萝卜素含量在Cr100和Cr200处理下分别比Cr0处理下降7.84%、25.49%。而在HA5和HA10处理下,植物胡萝卜素含量随Cr浓度增加下降显明显,其中在HA5处理下,植物胡萝卜素含量在Cr100和Cr200处理下分别比Cr0处理下降10.71%和66.07%;而HA10处理下,植物胡萝卜素含量在Cr100和Cr200处理下分别比Cr0处理下降 19.61% 和56.86%。在Cr100处理下,植物胡萝卜素含量随腐殖酸浓度的增加先增加后减少;而在Cr200处理下,植物胡萝卜素含量随腐殖酸浓度的增加先减少后增加。由此可以看出,腐殖酸处理在低浓度和高浓度Cr污染时会降低植物胡萝卜素含量。以上说明腐殖酸和Cr对植物胡萝卜素表现为协同作用。
2.3 重金属和腐殖酸对李氏禾叶绿素荧光参数的影响
叶绿素荧光参数能够直接或间接表征逆境中植物光合作用的光合机构、光合电子传递链及暗反应酶活性等生理生化指标受到的影响[15]。
Fo和Fm分别指光系统PSⅡ反应中心处于完全开放和完全关闭时的荧光产量[16]。从表3可以看出,在HA5和HA20处理下,植物初始荧光Fo和最大荧光Fm随着Cr浓度增加明显增加;HA5处理下,植物初始荧光Fo在Cr100和Cr200处理下分别比Cr0处理增加46.88%和 43.75%,而植物最大荧光Fm在Cr100和Cr200处理下分别比Cr0处理增加28.57%和35.34%;而在HA20处理下,植物最大荧光Fm在Cr100和Cr200处理下分别比Cr0处理增加21.88%和46.88%。在Cr0处理下,HA5、HA10处理植物初始荧光Fo和最大荧光Fm随着腐殖酸浓度的增加而逐渐增加;而在Cr100和Cr200处理下,植物初始荧光Fo和最大荧光Fm的变化不显著。
Fv/Fm常被用作表明环境胁迫程度的探针。Fv/Fm是 PSⅡ 最大光化学量子产量,开放的PSⅡ反应中心捕获激发能的效率,非胁迫条件下该参数的变化较小,一般植物恒定在 0.75~0.85,胁迫条件下该参数明显下降。从表3可以看出,在HA0处理下,PSⅡ最大光化学量子效率(Fv/Fm)随Cr浓度的升高无显著变化。在Cr0和Cr200处理下,PSⅡ最大光化学量子效率(Fv/Fm)随腐殖酸浓度的增加呈先增大后减小的趋势;而Cr100处理下,PSⅡ最大光化学量子效率(Fv/Fm)随腐殖酸浓度的增加先减少后增大。由此可以看出,腐殖酸在清洁土壤下PSⅡ最大光化学量子效率(Fv/Fm)降低,说明开放的PSⅡ反应中心捕获激发能的效率有所降低;而腐殖酸在高浓度Cr污染情况下,PSⅡ反应中心捕获激发能的效率随着腐殖酸浓度的增加,光合电子链的传递速率得到一定程度的恢复,从而维持正常的暗反应。
光化学淬灭系数(qp)值反映PSⅡ原初电子受体的氧化还原状态和PSⅡ开放中心的数量,其值越大,说明PSⅡ的电子传递活性越高;(qp)的下降表明PSⅡ反应中心的开放比例和参与CO2固定的能量减少[17]。非光化学淬灭系数(qN)值反映的是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的激发能部分,其值代表光合机构的损伤程度。如表3所示,同一浓度腐殖酸处理下,qN在Cr100和Cr200处理下较Cr0处理略有下降;而在HA20处理下,qp在Cr200处理下较Cr0处理明显升高,qN在Cr200处理下较Cr0处理明显下降。在Cr0和Cr100处理下,qp和qN随腐殖酸浓度的增大变化不大;而Cr200处理下,qp和qN随腐殖酸浓度的增大有所下降但差异不显著。由此可以看出,腐殖酸在清洁土壤和低浓度Cr污染时,qp和qN变化不大,而在高浓度Cr污染土壤情况下,qp和qN有所下降,表明叶片吸收光能的光化学反应减弱。
ETR是反映实际光照度条件下的表观电子传递速率。如表3所示,在同一浓度腐殖酸处理下,ETR随Cr浓度的增加先降低后升高。而在同一浓度Cr处理下,ETR随腐殖酸浓度的增加有所降低;尤其在同一浓度Cr200处理下,ETR随腐殖酸浓度的增加显著降低。由此可以看出,在Cr污染情况下可以影响光合电子传递速率ETR,说明在Cr污染土壤下ETR具有较高的损伤修复能力;而腐殖酸添加在清洁土壤和Cr污染土壤下会降低ETR,说明其表观电子传递受阻。
3 讨论
生物量是衡量植物耐性的指标,研究发现,适量的Cr能够促进植物的生长。在重金属胁迫下,植物的生物量可以作为评价植物对重金属耐性的临界指标。同时,生物量也可以直接影响重金属污染土壤的植物修复效果。有研究表明,六价铬[Cr(Ⅵ)]胁迫对白骨壤幼苗的萌发具有一定的促进作用,而高浓度则产生抑制作用[18]。本试验发现,在相同Cr100处理下李氏禾各项生长指标较Cr0明显增加,说明低浓度Cr对李氏禾生长有促进作用;腐殖酸在Cr污染土壤下,李氏禾的地上部干质量增加,说明低浓度Cr和腐殖酸能够促进李氏禾生长,这与适量的腐殖酸能够促进水稻地上部生长发育,表现为株高增加、生物量明显提高,这与对小麦、水稻等的研究[19]一致。
叶绿素在植物進行光合作用过程中起到接受和转换能量的作用,叶绿素含量与叶片衰老氧化密切相关,其变化必然导致植物光合作用的变化,从而影响植物的生长。叶绿素荧光与光合作用中各个反应过程紧密相连,任何逆境过程对光合作用各过程产生的影响通过体内叶绿素荧光诱导动力学参数变化反映出来[20]。鲁先文等研究表明,低浓度Cr促进小麦叶绿素的合成,高浓度Cr则产生毒害[21]。本试验中植物叶绿素含量随Cr含量增加呈现上升趋势,说明在Cr的刺激下,促进植物叶绿素的合成,增大了光合作用,影响李氏禾的生长,造成李氏禾的生物量增加,叶绿素含量变化趋势与鲜质量变化趋势相似[22]。而在清洁土壤和低浓度Cr污染土壤下中,腐殖酸的加入可以增加叶绿素含量,促进植物叶绿素生长。叶绿素在光合作用中起接受和转换能量的作用,叶绿素a/b值的降低说明其捕获和转化光能的能力下降。腐殖酸可以促进镁离子(Mg2+)和亚铁离子(Fe2+)的吸收,从而促进叶绿素含量增加[23]。叶绿素荧光变化反映植物光合机构的运转状况,荧光参数Fv/Fm的变化表明PSⅡ的原始光能转化效率能率[24]。本试验结果表明,低浓度的Cr促进李氏禾 Fv/Fm 值增加,但随着Cr浓度的增加,李氏禾Fv/Fm值呈下降趋势,这说明李氏禾高浓度Cr污染下降低了PSⅡ反应中心进行的光化学反应的效率。qp和qN反映了叶片对激发能利用情况,在相同Cr处理下,低浓度腐殖酸对李氏禾qp和qN影响不大,而在高浓度腐殖酸处理下,李氏禾qp和qN降低,说明植物叶片吸收光能的光化学反应减弱,而过剩光能通过非辐射能量(如热能)途径耗散受阻。
4 结论
李氏禾在低浓度Cr污染土壤下可以较好地生长。同时在Cr污染土壤下,李氏禾各部位干质量随腐殖酸浓度增高呈升高趋势。在200 mg/kg Cr污染土壤下,李氏禾没有表现出明显的毒害作用,腐殖酸的加入也能提高李氏禾地上部干质量。在清洁土壤和Cr污染土壤中,腐殖酸能够促进李氏禾叶绿素含量的,随腐殖酸浓度增高而升高。低浓度Cr和腐殖酸促进李氏禾光合生理的影响,并未出现抑制现象。
参考文献:
[1]邢艳帅,乔冬梅,朱桂芬,等. 土壤重金属污染及植物修复技术研究进展[J]. 中国农学通报,2014,30(17):218-224.
[2]申嘉澍,冯宇佳,王倩倩,等. 土壤重金属污染现状及其植物修复研究进展[J]. 绿色科技,2015,(12):226-227,229.
[3]郭修平. “土十条”与土壤污染治理[J]. 中华环境,2016,32(8):5.
[4]郭 凌,卜玉山,张 曼,等. 煤基腐殖酸对外源砷胁迫下玉米生长及生理性状的影响[J]. 环境工程学报,2014,8(2):758-766.
[5]Zhang X,Gao B,Xia H. Effect of cadmium on growth,photosynthesis,mineral nutrition and metal accumulation of bana grass and vetiver grass[J]. Ecotoxicology and Environmental Safety,2014,106:179-187.
[6]张学洪,罗亚平,黄海涛,等. 一种新发现的湿生铬超积累植物-李氏禾(Leersia hexandra Swartz)[J]. 生态学报,2006,26(3):950-953.
[7]丁文川,田秀美,王定勇,等. 腐殖酸对生物炭去除水中Cr(Ⅵ)的影响机制研究[J]. 环境科学,2012,33(11):3847-3853.
[8]钟桐生. 土壤腐殖酸性质及其化学传感器的研究[D]. 长沙:湖南大学,2009.
[9]闫双堆,卜玉山,刘利军,等. 不同腐殖酸物质对土壤中汞的固定作用及植物吸收的影响[J]. 环境科学学报,2007,27(1):101-105.
[10]Wang Q,Li Z,Cheng S,et al. Effects of humic acids on phytoextraction of Cu and Cd from sediment by Elodea nuttallii[J]. Chemosphere,2010,78(5):604-608.
[11]Vargas C,Pérez-Esteban J,Escolástico C,et al. Phytoremediation of Cu and Zn by vetiver grass in mine soils amended with humicacids[J]. Springer Berlin Heidelberg,2016,23(13):13521-13530.
[12]张杏锋,田 超,高 波. 能源植物皇草对重金属的耐性及修复潜力[J]. 环境工程学报,2017,11(5):3204-3213.
[13]陈全战,唐 宁,张边江,等. Cr(6+)对空心蓮子草光合生理特性的影响[J]. 湖北农业科学,2015,54(21):5310-5312,5317.
[14]余顺慧,方荣美,唐 洁,等. 铬污染对延胡索生长和生理特性的影响[J]. 江苏农业科学,2016,44(10):236-239.
[15]邵 云,王文斐,李学梅,等. 三种有机物料对铬污染土壤理化性质及小麦生长发育特征的影响[J]. 麦类作物学报,2016,36(6):779-788.
[16]罗小玲,李淑仪,蓝佩玲,等. 硅酸盐及腐植酸对Cr-Pb污染土壤中小白菜的生长和生理的影响[J]. 农业环境科学学报,2008,27(6):224-230.
[17]鲁 艳,李新荣,何明珠,等. 重金属对盐生草光合生理生长特性的影响[J]. 西北植物学报,2011,31(2):370-376.
[18]万永吉. 重金属Cr(Ⅵ)对红树植物白骨壤幼苗生长及生理生态效应的研究[D]. 厦门:厦门大学,2008.
[19]侯 明,张兴龙,路 畅,等. V(Ⅴ)、Cr(Ⅵ)单一和复合胁迫对小麦幼苗生长和生理特性的影响[J]. 环境化学,2012,31(7):1016-1022.
[20]钱永强,周晓星,韩 蕾,等. Cd2+胁迫对银芽柳PSⅡ叶绿素荧光光响应曲线的影响[J]. 生态学报,2011,31(20):6134-6142.
[21]鲁先文,余 林,宋小龙,等. 重金属铬对小麦叶绿素合成的影响[J]. 安徽农学通报,2007,13(14):101-102.
[22]周 武,袁志辉,刘敏超. 重金属Cr(Ⅵ)和Ni对旱伞草富集能力及其生理生化指标的影响[J]. 环境工程学报,2015,9(1):171-176.
[23]陈玉玲. 腐植酸对植物生理活动的影响[J]. 植物学通报,2000,17(1):64-72.
[24]王碧霞,肖 娟,冯 旭,等. 铬胁迫对葎草雌雄植株光合生理特性的不同影响[J]. 草业学报,2016,25(7):131-139.张洪宇,张 晶,吴建寨,等. 乡村治理科技2050发展战略研究[J]. 江苏农业科学,2019,47(5):268-272.
- 供给侧改革背景下我国城市马拉松发展路径研究
- 不同负荷游泳运动对大鼠部分免疫指标的影响
- 近6届全国冬季运动会自由式滑雪空中技巧比赛的对比分析
- 我国优秀单板滑雪U型场地技巧运动员心理健康状况的调查研究
- 体育教育专业学生基本功大赛开展的现状及对策研究
- 电子类游戏对少年儿童动商的影响及体育游戏干预
- 老年人体育需求及社会支持体系研究
- 太极拳人才培养的机遇、挑战与思考
- 独竹漂项目衍变历程对我国民族传统体育发展的启示
- 体育图腾及其崇拜的表现形式与社会效应研究
- 公共体育信息服务研究:轨迹、诊断与启示
- 中心动脉压、增益指数与年龄、性别相关性研究
- 体育运动参与程度与大学生人格特征和社会适应能力的关系研究
- 基于知识图谱的国际高强度间歇训练研究可视化分析
- 世界大学生三对三篮球联赛传切配合分析
- 吉林省警察院校学生体能训练现状与对策研究
- 高校休闲潜水课程中的生命教育
- 高校体育英语专业《现代汉语》教学的困境与思考
- 海南省体育场地建设现状与发展对策
- 山东省民俗民问体育活动的发展特征与区域特色研究
- 宁夏政府购买公共体育服务现状研究
- 上海国际大型体育赛事本土化研究
- 国家级非物质文化遗产佛汉拳研究综述
- 东北三省冬季传统体育消费者现状的调查研究
- 社会化CRM理论在职业足球俱乐部营销中的应用研究
- prestructuring
- prestruggle
- prestruggled
- prestruggles
- prestruggling
- prestubborn
- prestudied
- prestudies
- prestudious
- prestudiously
- prestudiousness
- prestudiousnesses
- prestudy
- prestudying
- presubdue
- presubdued
- presubdues
- presubduing
- presubject
- presubjected
- presubjecting
- presubjects
- presubmission
- presubmissions
- presubmit
- 心思灵巧,很会说话
- 心思灵敏
- 心思灵敏,手艺工巧
- 心思灵活
- 心思相同
- 心思精力
- 心思纷乱纠结如茧
- 心思细密
- 心思翻腾,内心很不平静
- 心思,心情
- 心思;主意;想法
- 心怡神悦
- 心怡神旷
- 心急
- 心急只恨马行慢
- 心急吃不上热馒头
- 心急吃不了热包子
- 心急吃不了热年糕
- 心急吃不了热汤面
- 心急吃不了热豆腐
- 心急吃不得热粥
- 心急吃热粥
- 心急如火
- 心急如焚
- 心急嫁不了好汉子