陈邦兰 翟梦倩 龙涛 赵顺程 刘娟 尹茂灵 万心言 王治杰 刘继恺


摘要?自然抗性相關巨噬细胞蛋白(natural?resistance?associated?macrophage?proteins,NRAMPs)是一类高度保守的二价金属跨膜转运蛋白,在调控生物体内重金属稳态中发挥着重要作用。为了探究烟草NRAMP基因的多样性,该研究根据烟草基因组数据信息,利用生物信息学方法对烟草NRAMP基因家族成员进行了鉴定,对其基本信息、结构特点、蛋白理化性质及保守序列、启动子元件和组织表达模式等进行了分析。结果表明:烟草基因组含有9个NtNRAMP基因,分别命名为NtNRAMP1.1~6.2,其编码氨基酸长度为500~542?aa,蛋白分子量为54.31~59.11?kD,等电点为5.09~8.73,总平均亲水性为0.475~0.604;除了NRAMP6.1和6.2不含有motif4外,其余NRAMP蛋白都含有motif1~motif10。顺式作用元件分析表明所有NtNRAMP基因启动子中均含有参与基因转录、非生物胁迫、植物激素响应和光应答途径的元件,部分基因含有生物胁迫、次生代谢、组织表达和结合位点的元件。染色体定位发现,6个NtNRAMP基因分别分布在5条烟草染色体上。表达分析表明:NtNRAMP基因组织表达模式差异较大,其中NtNRAMP6.1和NtNRAMP6.2基因表达具有组织特异性。该研究结果为进一步研究烟草NtNRAMP基因的功能奠定了基础。
关键词?自然抗性相关巨噬细胞蛋白;烟草;基因家族;表达分析
中图分类号?S572?文献标识码?A?文章编号?0517-6611(2021)03-0100-05
doi:10.3969/j.issn.0517-6611.2021.03.027
Abstract?Natural?resistance?associated?macrophage?proteins?(NRAMPs)?are?highly?conserved?bivalent?metal?transporters?that?play?an?important?role?in?regulating?heavy?metal?homeostasis.?In?order?to?explore?the?diversity?of?the?NRAMP?gene?in?tobacco,?this?study?identified?the?members?of?the?NRAMP?gene?family?of?tobacco?by?using?bioinformatics?methods?based?on?the?genome?data?of?N.?tabacum,?and?analyzed?the?basic?information,?structural?features,?physicochemical?parameters,?conserved?motifs,?cisacting?elements,?tissue?expression?pattern?of?these?NtNRAMP?genes.?The?results?showed?that?N.?tabacum?genome?contained?9?NtNRAMP?genes?which?were?designated?as?NtNRAMP1.16.2.?The?length?of?NtNRAMP?proteins?ranged?from?500?to?542?amino?acid;the?relative?molecular?weight?ranged?from?54.31?to?59.11?kD;the?predicted?pI?ranged?from?5.09?to?8.73,?and?the?predicted?GRAVY?ranged?from?0.475?to?0.604.?Except?for?NRAMP6.1?and?NRAMP?6.2,?which?did?not?contained?motif4,?all?NtNRAMP?proteins?contained?motif1motif10.?Cisacting?regulatory?element?prediction?showed?that?all?NtNRAMP?genes?contained?elements?involved?in?gene?transcription,?abiotic?stress,?plant?hormone?response?and?light?response?pathways,?however,?some?genes?contained?elements?involved?in?biological?stress,?secondary?metabolism,?tissue?expression?and?binding?sites.?Chromosomal?localization?revealed?that?6?of?9?NtNRAMP?genes?were?located?in?5?tobacco?chromosomes.?The?tissue?expression?profile?analysis?showed?diverse?tissue?expression?patterns?of?NtNRAMPs,?and?NtNRAMP6.1?and?NtNRAMP6.2?showed?tissue?specific?expression?patterns.?These?results?laid?a?foundation?for?further?functional?characterization?of?NtNRAMP?genes.
Key?words?Natural?resistance?associated?macrophage?proteins;Nicotiana?tabacum;Gene?family;Expression?analysis
Cu、Fe、Mn和Zn等过渡金属是生物体重要的微量元素,在植物生长发育和新陈代谢中起着关键作用[1]。但同时,这些金属含量又必须保持在合理水平,一旦过量则会对细胞产生毒害。还有一些金属,如Cd、Pd和Hg等,它们虽然不是必需元素,但也可以被植物吸收,对植物造成负面影响[2]。为了适应环境变化,植物在长期进化过程中形成了一套复杂且精密的调控过程以保证体内重金属的稳态,而重金属转运蛋白在这一调控过程中发挥着关键作用[3]。
自然抗性相关巨噬细胞蛋白(natural?resistance?associated?macrophage?proteins,NRAMPs)是一类高度保守的二价金属跨膜转运蛋白,广泛存在于细菌、酵母、果蝇、人类和植物中[4]。NRAMP基因最早在老鼠中被克隆,该基因在老鼠抵御分枝杆菌过程中起到重要作用[5]。目前,科学家们已克隆到多个植物NRAMP基因,并对部分NRAMP基因进行了功能研究。模式植物拟南芥有6个NRAMP基因(AtNRAMP1~6)。已有研究发现,AtNRAMP1是定位于拟南芥根细胞质膜上的高亲和性Mn转运蛋白,该蛋白负责拟南芥在低Mn环境对Mn的吸收[6]。AtNRAMP2是一種定位于反面高尔基网络(TransGolgi?Network)的Mn转运蛋白,该蛋白在维持细胞内Mn的正常分布以及Mn缺乏条件下根的生长中发挥重要作用[7-8]。与AtNRAMP1和AtNRAMP2不同,AtNRAMP3和AtNRAMP4蛋白定位于液泡膜上,是Fe和Mn的转运蛋白,调控Fe缺乏下的种子萌发以及Mn缺乏下植物的光合作用和生长[9-11]。AtNRAMP6定位于拟南芥侧根和幼嫩叶片细胞的高尔基体或反面高尔基网络上,是Cd和Fe的转运蛋白[12-13]。
烟草是我国重要的非食品性经济作物,易吸收和富集重金属,这些重金属可通过吸烟进入人体,危害人体健康[14]。对烟草金属转运蛋白进行鉴定和功能研究对于进一步阐明烟草对重金属的吸收富集机制,保证烟叶安全性是十分必要的。之前在烟草中鉴定到26个金属耐受蛋白(metal?tolerance?protein,MTP)编码基因,并发现NtMTP?8.1、NtMTP8.4和NtMTP11.1可以在酵母中转运Mn[15]。该研究采用生物信息学手段对烟草NRAMP基因家族进行鉴定,并对它们的基因特征和组织表达模式进行分析,旨在为烟草NRAMP基因的功能研究奠定基础。
1?材料与方法
1.1?烟草NRAMP基因的鉴定
以拟南芥中6个NRAMP蛋白氨基酸序列作为查询序列,在烟草基因组数据库(Nitab?v4.5?cDNA?Edwards,2017,https:∥solgenomics.net/)中进行TBLASTN比对搜索。去除冗余序列后,将得到的序列用InterProScan(http:∥www.ebi.ac.uk/interpro/)工具验证是否属于NRAMP家族(IPR001046)[16]。经验证后的序列为烟草NtNRAMP蛋白序列,下载对应的基因号、基因组位置等信息。将获得的NtNRAMP蛋白序列在NCBI中分别与拟南芥6个AtNRAMP蛋白序列作比对,综合序列一致性和覆盖度结果对NtNRAMP基因进行命名。
1.2?烟草NRAMP蛋白生化参数和结构特征
利用ProParam工具预测烟草NRAMP蛋白的分子量、等电点和亲水性[17],利用PlantmPloc预测亚细胞定位[18],利用TMHMM?Server?V.2.0预测跨膜结构域[19-20],利用MEME程序预测保守基序(motif)[21]。
1.3?烟草NRAMP基因结构和染色体定位分析
利用Sol?Genomics?Network(SGN)网站(https:∥solgenomics.net/)烟草基因组注释信息分析烟草NRAMP基因结构和染色体定位。最后,运用TBtools软件进行外显子/内含子结构、蛋白结构和染色体定位作图[22]。
1.4?烟草NRAMP基因顺式作用元件及组织表达特性分析
在上述SGN网站的烟草基因组数据库(Nitab?v4.5?Genome?Scaffolds?Edwards,2017)中搜索获得烟草NRAMP基因上游1?500?bp序列,利用在线网站PlantCARE对获得的序列进行顺式作用元件的预测[23]。为了研究烟草NRAMP基因的组织表达模式,从GenBank?Sequence?Read?Archive(SRA)上下载烟草TN90的RNA转录组数据(登记号SRP029183),分析目的基因在8个不同烟草组织中的相对表达量并用TBtools软件作图。
2?结果与分析
2.1?烟草NRAMP基因的鉴定及蛋白理化性质分析
利用生物信息学方法在烟草基因组数据库中进行检索,共鉴定出9个NtNRAMP基因,综合这9个基因编码蛋白序列与拟南芥AtNRAMP序列的比对结果,分别将它们命名为NtNRAMP1.1、1.2、2.1、2.2、2.3、3.1、3.2、6.1、6.2(表1)。
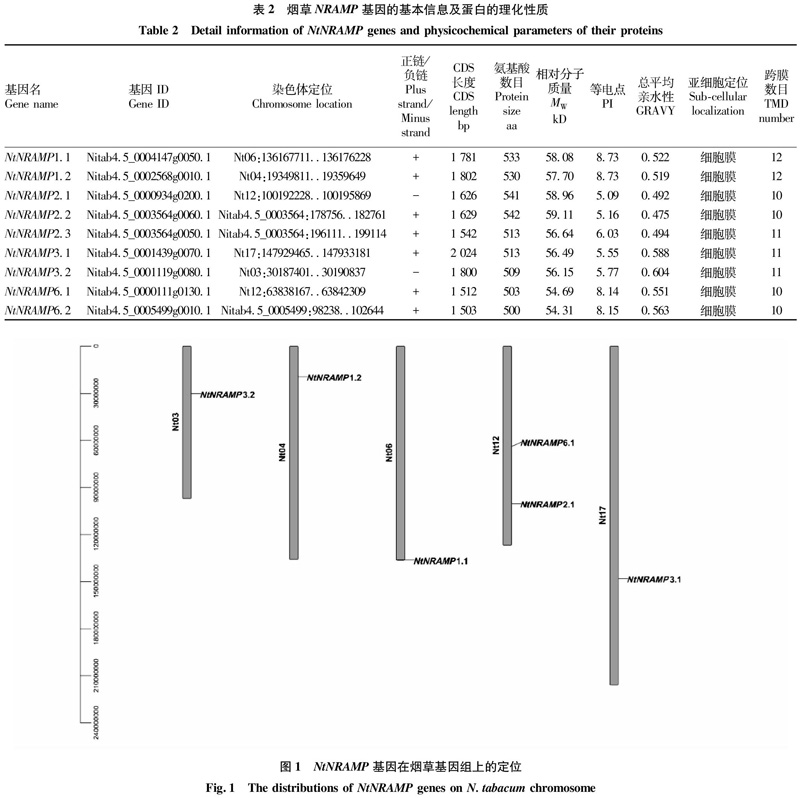
利用在线软件对NtNRAMP基因及其编码蛋白进行理化特性分析,结果表明(表2),NtNRAMP基因蛋白质编码区(coding?sequence,CDS)长度在1?503?bp(NtNRAMP6.2)~2?024?bp(NtNRAMP3.1),平均为1?691?bp;氨基酸个数在500?aa(NtNRAMP6.2)~542?aa(NtNRAMP2.2);相对分子质量介于54.31~59.11?kD;等电点为5.09~8.73,其中NRAMP1.1、1.2、6.1、6.2为碱性蛋白,其余为酸性蛋白;总平均亲水性为0.475~0.604;跨膜结构域(transmembrane?domain,TMD)数目相近,NRAMP2.1、2.2、6.1、6.2有10个TMD,NRAMP2.3、3.1和3.2有11个TMD,NRAMP1.1和1.2有12个TMD;值得注意的是,9个NtNRAMP蛋白均被预测定位于细胞膜。
2.2?烟草NRAMP基因的染色体分布
对NtNRAMP基因的染色体定位进行分析,结果显示(图1),6个NtNRAMP基因分布于5条烟草染色体上(分别是3、4、6、12、17号染色体),其中12号染色体含有2个NtNRAMP基因(NtNRAMP2.1和NtNRAMP6.1),其余4条染色体各含有1个NtNRAMP基因。
2.3?烟草NRAMP基因结构和蛋白质保守序列分析
利用NtNRAMP基因的注释信息和TBtools软件分析烟草NRAMP的基因结构。结果如图2所示,NRAMP2.1、2.2、2.3、3.1、3.2基因具有相同的外显子数目(4个),NRAMP6.1和6.2基因具有12个外显子,NRAMP1.1和1.2基因具有13个外显子,其中NRAMP1.1、1.2、3.1、3.2基因存在上游和下游非翻译区(UTR)。蛋白质保守序列分析结果显示:除了NRAMP6.1和6.2不含有motif4外,所有NRAMP蛋白都含有motif1~motif10。
2.4?烟草NRAMP基因家族顺式作用元件分析
通过分别对9个NtNRAMP基因上游1?500?bp序列进行分析,预测其可能含有的顺式作用元件。结果显示(表3),所有NtNRAMP基因均含有参与基因转录、非生物胁迫、植物激素响应、光应答途径的元件,其中含有涉及基因转录的作用元件最多(45~72个);此外,NRAMP1.1和1.2还各含有1个生物胁迫元件,NRAMP3.2、6.1、6.2各含有1个次生代谢元件,除NRAMP2.3和3.1外,其余基因启动子中均含有1~2个组织表达元件,除NRAMP1.1、2.3、3.2外,其余基因启动子中均含有1~2个结合位点元件。
2.5?烟草NRAMP基因组织表达模式分析
利用转录组数据分析烟草NRAMP基因分别在幼嫩/成熟的花、幼嫩/成熟/衰老的叶、干燥的蒴果、根、茎8个组织中的表达情况。结果如图3所示,NRAMP1.1基因的表达在整体上均高于NRAMP1.2,NRAMP1.1基因在根中的表达量最高,而NRAMP1.2基因在茎中表达量最高,在干燥的蒴果里几乎不表达;NRAMP2.1和2.2在烟草各组织中均有表达,具有相似的组成型表达模式;NRAMP2.3与NRAMP2.1和2.2相比,在各组织中的表达量相对较低,特别在根、干燥的蒴果、成熟/衰老的叶中表达量很低;NRAMP3.1和3.2基因的组织表达模式相似,在烟草各组织中均有表达,且在幼嫩的叶中表达量最高;NRAMP6.1和NRAMP6.2基因的表达具有组织特异性,NRAMP6.1基因只在根和成熟的花中表达,而NRAMP6.2基因还在幼嫩的花中表达,这2个基因在其余组织中均无表达或表达量极低。
3?讨论
金属转运蛋白,比如NRAMP,是生物体金属特异转运系统的主要组成部分,在维持细胞及其细胞器内重金属稳态方面发挥着重要作用。近年来,随着植物基因组学研究的不断深入,研究者们利用比较基因组学等方法成功对水稻、大豆和可可中的NRAMP基因家族进行了鉴定分析[24-26]。该研究利用同样的方法在烟草基因组中鉴定到9个NtNRAMP基因,同时运用生物信息学方法对这些基因的基本信息、结构特点、蛋白理化性质及保守序列、启动子元件和组织表达模式等进行了分析预测,这些结果将为烟草NRAMP基因家族功能研究以及其分子调控机制的阐明提供理论依据。
水稻含有7个NRAMP基因,这些基因编码蛋白的分子量在55.81~59.71?kD,含有10~12个跨膜结构域,且全部定位于质膜上[25]。大豆基因组包含13个NRAMP,它们的分子量在55.44~64.39?kD,GmNRAMP1a~GmNRAMP4b定位于液泡膜,GmNRAMP5a~GMNRAMP7定位于质膜[24]。拟南芥有6个NRAMP,它们分别在质膜、液泡膜和高尔基体上发挥作用。研究中,烟草NtNRAMP的分子量为54.31~59.11?kD,跨膜結构域为10~12,定位于细胞膜。通过比较这些结果可以发现,不同植物中NRAMP蛋白的分子量大小和跨膜结构域数目比较相似,但在亚细胞定位方面却存在差异。亚细胞定位是阐释金属转运蛋白生物学功能的重要数据,因此下一步有必要利用试验技术,如荧光蛋白表达结合共聚焦显微镜观察、免疫印迹等,对烟草NRAMP各成员的亚细胞定位进行深入研究。
为了探索调控NtNRAMP基因表达的潜在机制,该研究对烟草NtNRAMP基因上游的顺式作用元件进行了分析,共鉴定到656个顺式作用元件。其中,所有NtNRAMP基因启动子中都含有参与非生物胁迫、植物激素响应、光应答途径的顺式作用元件,表明NtNRAMP基因广泛参与到这些生物过程中。值得注意的是,和其他NRAMP基因相比,仅NRAMP1.1和1.2各含有1个生物胁迫元件,NRAMP3.2、6.1、6.2各含有1个次生代谢元件,说明这些基因还可能特异性地在生物胁迫响应和次生代谢途径中发挥作用。
烟草NRAMP基因在幼嫩/成熟的花、幼嫩/成熟/衰老的叶、干燥的蒴果、根、茎8个组织中有不同程度的表达,但组织表达模式差异较大,其中NtNRAMP6.1和NtNRAMP6.2基因只在根和花中表达,其表达具有组织特异性。最近的研究发现拟南芥AtNRAMP6基因能够调控Fe缺乏情况下拟南芥侧根的生长,而对主根没有影响[13]。因此,NtNRAMP6.1和NtNRAMP6.2可能会通过调控根和花细胞中金属稳态从而影响烟草根和花的生长发育。
参考文献
[1] HNSCH?R,MENDEL?R?R.Physiological?functions?of?mineral?micronutrients(Cu,Zn,Mn,Fe,Ni,Mo,B,Cl)[J].Curr?Opin?Plant?Biol,2009,12(3):259-266.
[2]?CLEMENS?S.Molecular?mechanisms?of?plant?metal?tolerance?and?homeostasis[J].Planta,2001,212(4):475-486.
[3]?金楓,王翠,林海建,等.植物重金属转运蛋白研究进展[J].应用生态学报,2010,21(7):1875-1882.
[4]?NEVO?Y,NELSON?N.The?NRAMP?family?of?metalion?transporters[J].Biochim?Biophys?Acta,2006,1763(7):609-620.
[5]?VIDAL?S?M,MALO?D,VOGAN?K,et?al.Natural?resistance?to?infection?with?intracellular?parasites:Isolation?of?a?candidate?for?Bcg[J].Cell,1993,73(3):469-485.
[6]?CAILLIATTE?R,SCHIKORA?A,BRIAT?J?F,et?al.Highaffinity?manganese?uptake?by?the?metal?transporter?NRAMP1?is?essential?for?Arabidopsis?growth?in?low?manganese?conditions[J].Plant?Cell,2010,22(3):904-917.
[7]?ALEJANDRO?S,CAILLIATTE?R,ALCON?C,et?al.Intracellular?distribution?of?manganese?by?the?transGolgi?network?transporter?NRAMP2?is?critical?for?photosynthesis?and?cellular?redox?homeostasis[J].Plant?Cell,2017,29(12):3068-3084.
[8]?GAO?H?L,XIE?W?X,YANG?C?H,et?al.NRAMP2,a?transGolgi?networklocalized?manganese?transporter,is?required?for?Arabidopsis?root?growth?under?manganese?deficiency[J].New?Phytol,2018,217(1):179-193.
[9]?THOMINE?S,LELIVRE?F,DEBARBIEUX?E,et?al.AtNRAMP3,a?multispecific?vacuolar?metal?transporter?involved?in?plant?responses?to?iron?deficiency[J].Plant?J,2003,34(5):685-695.
[10]?LANQUAR?V,LELIVRE?F,BOLTE?S,et?al.Mobilization?of?vacuolar?iron?by?AtNRAMP3?and?AtNRAMP4?is?essential?for?seed?germination?on?low?iron[J].EMBO?J,2005?24(23):4041-4051.
[11]?LANQUAR?V,RAMOS?M?S,LELIVRE?F,et?al.Export?of?vacuolar?manganese?by?AtNRAMP3?and?AtNRAMP4?is?required?for?optimal?photosynthesis?and?growth?under?manganese?deficiency[J].Plant?Physiol,2010,152(4):1986-1999.
[12]?CAILLIATTE?R,LAPEYRE?B,BRIAT?J?F,et?al.The?NRAMP6?metal?transporter?contributes?to?cadmium?toxicity[J].Biochem?J,2009,422(2):217-228.
[13]?LI?J?Y,WANG?Y?R,ZHENG?L,et?al.The?intracellular?transporter?AtNRAMP6?is?involved?in?Fe?homeostasis?in?Arabidopsis[J].Front?Plant?Sci,2009,10:1-11.
[14]?郎思曼,王龍宪,许自成,等.烟草对重金属的吸收分布特征及影响因素研究综述[J].江西农业学报,2012,24(11):93-99.
[15]?LIU?J?K,GAO?Y?F,TANG?Y?L,et?al.Genomewide?identification,comprehensive?gene?feature,evolution,and?expression?analysis?of?plant?metal?tolerance?proteins?in?tobacco?under?heavy?metal?toxicity[J].Front?Genet,2019,10:345.
[16]?FINN?R?D,ATTWOOD?T?K,BABBITT?P?C,et?al.InterPro?in?2017beyond?protein?family?and?domain?annotations[J].Nucleic?Acids?Res,2017,45(D1):D190-D199.
[17]?GASTEIGER?E,HOOGLAND?C,GATTIKER?A,et?al.Protein?identification?and?analysis?tools?on?the?ExPASy?server[M]∥WALKER?J?M.The?proteomics?protocols?handbook.Totowa,NJ:Humana?Press,2005:571-607.
[18]?CHOU?K?C,SHEN?H?B.PlantmPLoc:A?topdown?strategy?to?augment?the?power?for?predicting?plant?protein?subcellular?localization[J].PLoS?One,2010,5(6):1-11.
[19]?SONNHAMMER?E?L?L,VON?HEIJNE?G,KROGH?A.A?hidden?Markov?model?for?predicting?transmembrane?helices?in?protein?sequences[J].Proc?Int?Conf?Intell?Syst?Mol?Biol,1998,6:175-182.
[20]?CHANDRASEKARAN?J,BRUMIN?M,WOLF?D,et?al.Development?of?broad?virus?resistance?in?nontransgenic?cucumber?using?CRISPR/Cas9?technology[J].Molecular?plant?pathology,2016,17(7):1140-1153.
[21]?BAILEY?T?L,BODEN?M,BUSKE?F?A,et?al.MEME?SUITE:Tools?for?motif?discovery?and?searching[J].Nucleic?Acids?Res,2009,37(S2):W202-208.
[22]?CHEN?C?J,XIA?R,CHEN?H,et?al.TBtools,a?Toolkit?for?Biologists?integrating?various?HTSdata?handling?tools?with?a?userfriendly?interface[J/OL].bioRxiv,2018[2020-03-07].https:∥doi.org/10.1101/289660.
[23]?ROMBAUTS?S,DHAIS?P,VAN?MONTAGU?M,et?al.PlantCARE,a?plant?cisacting?regulatory?element?database[J].Nucleic?Acids?Res,1999,27(1):295-296.
[24]?QIN?L,HAN?P?P,CHEN?L?Y,et?al.Genomewide?identification?and?expression?analysis?of?NRAMP?family?genes?in?soybean(Glycine?max?L.)[J].Frontiers?in?plant?science,2017,8:1436-1455.
[25]?MANI?A,SANKARANARAYANAN?K.In?silico?analysis?of?natural?resistanceassociated?macrophage?protein(NRAMP)family?of?transporters?in?rice[J].Protein?J,2018,37(3):237-247.
[26]?ULLAH?I,WANG?Y?R,EIDE?D?J,et?al.Evolution,and?functional?analysis?of?Natural?ResistanceAssociated?Macrophage?Proteins(NRAMPs)from?Theobroma?cacao?and?their?role?in?cadmium?accumulation[J].Sci?Rep,2018,8:14412-14427.
- 支气管扩张患者行雾化吸入治疗的护理效果分析
- 护理干预对丙型病毒性肝炎患者抗病毒治疗依从性的作用分析
- 优质护理在妊高症产妇产后出血护理中的应用效果
- 儿科护士工作满意度与社会支持的相关性分析
- 优质护理在子宫肌瘤患者护理中的应用效果观察
- 心脏瓣膜置换术后微量泵高浓度补钾的临床护理体会
- 骨科病人营养干预对术后恢复的影响
- 疼痛护理在痔疮手术患者中的应用研究
- 伤寒观察及护理
- 急诊科护理干预对老年重症心力衰竭患者治疗效果的作用分析
- 护理措施对肺癌全肺切除术后并发症的影响分析
- 系统性护理干预措施对妊娠期高血压疾病患者影响分析
- I例肺癌合并下肢血栓性静脉炎的护理体会
- 持续护理干预对雾化吸入治疗小儿哮喘过程的影响
- 人性化护理在75例剖宫产产科护理的效果观察
- 针灸按摩配合综合康复护理对高血压脑出血后肢体偏瘫的影响
- 优质护理模式在妊娠期糖尿病合并妊娠期高血压产妇中的应用
- 人性化服务在麻醉护理中的应用分析
- 护理风险管理在肿瘤患者护理中的应用
- 全面护理方式对小儿癫痫护理工作的影响分析
- 外周静脉置入中心静脉导管在肿瘤护理中的应用价值
- 关于儿童预防接种的护理体会
- 个体化护理应用在胆结石手术护理中的临床效果观察
- 悬浮牵引治疗胸腰椎压缩骨折的护理
- 社区护理干预对青少年近视及眼部感染防控效果观察
- set sb back sth
- set sb free
- set sb/sth apart
- set sb/sth back
- set sb/sth free
- set sb/sth straight
- set sb straight
- set sb up
- sets forth
- sets in
- set somebody off (doing something)
- set somethingforth
- set something in/into something
- set somethingoff
- set somethingout
- sets out
- set square
- set squares
- set sth against sth
- set sth aside
- set sth down
- set sth forth
- set sth going
- set sth in motion / put sth in motion
- set sth off
- 关门状
- 关门的
- 关门落闩
- 关门踩高跷——只知道自己高
- 关门踩高跷——自看自高
- 关门过日子
- 关门閤户
- 关门闭户
- 关门防客走
- 关门骂皇帝
- 关闭
- 关闭停业
- 关闭大门,安分守已
- 关闭铺子,停止营业
- 关闭门户,不外出与人交往接触
- 关闭限制
- 关闭,使隔绝
- 关阃
- 关防
- 关防严密
- 关防盆儿
- 关陆
- 关限
- 关隔
- 关隘