余小珍 黄维清 孙鹏鹏 阮迎春 刘志晶 陈桦

[摘要]目的分析过表达OCT4后人毛囊干细胞中转录本的变化及OCT4靶基因的表达。方法分别收集OCT4转导前后的毛囊干细胞并提取总RNA,利用转录组测序分析全基因转录本的差异,并分析差异表达基因(DEGs)的功能以及OCT4靶基因的表达。结果转导OCT4后的贴壁细胞和悬浮细胞亚群中分别有4 283和6 206个DEGs,转录本发生显著改变。上调DEGs显著富集于中胚层和外胚层发育的生物过程中,以及调节干细胞多潜能性的信号通路中。多潜能性相关的OCT4靶基因表达显著上调,胚胎造血基因KDR表达水平持续上调。结论OCT4转导后人毛囊干细胞的转录组发生显著改变并获得更多分化潜能,且悬浮细胞亚群呈现造血分化倾向。
[关键词]OCT4;毛囊;多潜能干细胞;细胞重新编程;造血干细胞
[中图分类号]R329.25[文献标志码]A[文章编号]2096-5532(2021)01-0077-05
[ABSTRACT]ObjectiveTo investigate the change in transcripts and the expression of the target genes of OCT4 in human hair follicle stem cells after OCT4 overexpression. MethodsTotal RNA was extracted from hair follicle stem cells before and after OCT4 transduction, and transcriptome sequencing was used to analyze the difference in transcripts. The function of differentially expressed genes (DEGs) and the expression of the target genes of OCT4 were analyzed. ResultsA total of 4 283 and 6 206 DEGs were screened out in the adherent cells and suspension cells, respectively, after OCT4 transduction, and there was a significant change in transcripts. Upregulated DEGs were mainly enriched in the biological processes of mesoderm and ectoderm development and the signaling pathways involved in regulating pluripotency of stem cells. The target genes of OCT4 associated with pluripotency were significantly upregulated, and the expression of the embryonic hematopoietic gene KDR continued to increase. ConclusionAfter OCT4 transduction, human hair follicle stem cells show a significant change in transcriptome and obtain a higher level of pluripotency, and the suspension cells show a tendency of hematopoietic differentiation.
[KEY WORDS]OCT4; hair follicle; pluripotent stem cells; cellular reprogramming; hematopoietic stem cells
利用基因工程技術能够将一种细胞重编程并直接转分化成另一种细胞[1-2]。我们的前期研究将这一技术应用于人毛囊干细胞(hHFMSC),并成功地将其转分化为红细胞。过表达OCT4的hHFMSC在培养过程中从贴壁细胞中逐渐产生一类小而圆、易悬浮的新的细胞亚群,该细胞亚群经造血诱导培养和红系生长因子刺激后能够向成熟的红细胞转分化[3]。hHFMSC本身具有多向分化的潜能[4],在转导4个多潜能转录因子(OCT4、SOX2、KLF4和c-Myc)后可以被重编程为诱导多能干细胞(iPSC),而单独转导OCT4可以被诱导生成红细胞,但OCT4重编程hHFMSC的机制尚未有研究。本研究在前期研究的细胞模型基础上,利用转录组测序分析OCT4转导后全基因转录组的改变,通过功能富集分析以及KEGG分析探索OCT4对各谱系细胞发育的核心调控作用,筛查OCT4下游靶基因,旨在探讨OCT4重编程hHFMSC的分子机制。
1材料与方法
1.1细胞样本及培养方法
前期工作中分离并鉴定后获得的hHFMSC;hHFMSCOCT4-adherent:应用含有OCT4 cDNA的慢病毒载体pLV-EF1α-OCT4-IRES-EGFP以及含有表达pVSVG、gag-pol和rev包装质粒共同转导hHFMSC,并经OCT4基因表达鉴定的贴壁细胞;hHFMSCOCT4-floating:收集hHFMSCOCT4-adherent的上层培养液,离心重悬后获得的悬浮细胞。
培养条件:hHFMSC接种于培养瓶或培养皿中,hHFMSCOCT4-adherent、hHFMSCOCT4-floating细胞接种于Matrigel包被的培养板上。3种细胞均用含体积分数0.10胎牛血清、10 μg/L碱性成纤维细胞生长因子(bFGF)的H-DMEM/F12培养液,置于37 ℃、含体积分数0.05 CO2培养箱中恒温培养。
1.2RNA提取及测序
每组细胞准備3个重复的独立样本,送至上海欧易公司进行转录组测序。按照说明书的操作步骤,使用mirVana miRNA Isolation Kit提取各样本总RNA,应用Agilent 2100 Bioanalyzer进行质检,将RNA完整性编号(RIN)≥7的样本被纳入后续分析。富集的mRNA加入打断试剂使其片段化,合成的双链cDNA经纯化、修复和连接测序接头后进行PCR扩增,应用TruSeq Stranded mRNA LTSample Prep Kit构建文库,通过Illumina HiSeq X Ten sequencer进行测序。
1.3差异表达基因(DEGs)分析及富集计算
使用Trimmomatic软件对Raw reads进行质控,将经去接头、低质量数据和碱基过滤处理得到的Clean reads与参考基因组进行比对,使用cufflinks和DESeq软件分别对基因表达进行定量和标准化,按照差异倍数(fold change,FC)>2或<-2以及P<0.05的标准筛选出DEGs。利用DAVID数据库中的Function Annotation 工具对DEGs分别进行GO功能富集和KEGG信号通路富集分析。
1.4统计学分析
利用R package计算Pearson相关系数及进行负二项分布检验。相关系数越接近1,表示样本间的相似度越高;负二项分布检验用于拟合基因覆盖的Reads数、校正由基因长度引起的误差和对基因表达量的差异进行假设检验。以P<0.05为差异具有统计学意义。
2结果
2.1基因转录本的差异分析
转录组测序结果显示,与hHFMSC相比较,hHFMSCOCT4-adherent中共有4 283个DEGs,其中上调者2 401个,下调者1 882个;hHFMSCOCT4-floating中有6 106个DEGs,其中上调基因3 107个,下调基因2 999个;与 hHFMSCOCT4-adherent相比,hHFMSCOCT4-floating中筛选出1 940个DEGs,其中上调基因833个,下调基因1 107个。见图1。表明各组细胞样本中基因组转录本具有显著差异,且悬浮细胞与hHFMSC相比DEGs数目最多;另外,同为OCT4转导后的细胞,悬浮细胞与贴壁细胞亚群的转录本明显不同,说明二者可能具有不同的分化潜能。
2.2DEGs的聚类分析
根据每组样本DEGs的表达量绘制热图(图2)进行水平聚类分析,结果显示,各组内样本水平聚类距离更近,而组间样本聚类距离相对较远,表明各组内样本具有良好的重复性,组间样本的基因表达量具有显著差异。
2.3DEGs的功能富集分析
使用DAVID数据库分别对生物过程、细胞组分和分子功能进行富集分析的结果显示,贴壁细胞和悬浮细胞分别与hHFMSC相比,上调基因显著富集于中胚层(胸腺、肢体、血管、肌肉、骨骼、心脏)及外胚层(内耳、脑、神经系统、皮肤、乳房)的器官发育及细胞分化生物过程中(图3)。表明hHFMSC在转导OCT4后获得了更多方向的分化潜能,甚至可能具有跨胚层发育的潜能。使用DAVID数据库对KEGG信号通路进行富集分析的结果显示,贴壁细胞和悬浮细胞分别与hHFMSC相比,调节干细胞多潜能性的信号通路中分别有29个和31个基因表达上调,包括ID3、BMP4、KLF4、LEFTY2和MYC等。进一步说明转导OCT4可能通过该通路调节hHFMSC的多潜能性。见表1。
2.4OCT4靶基因的表达
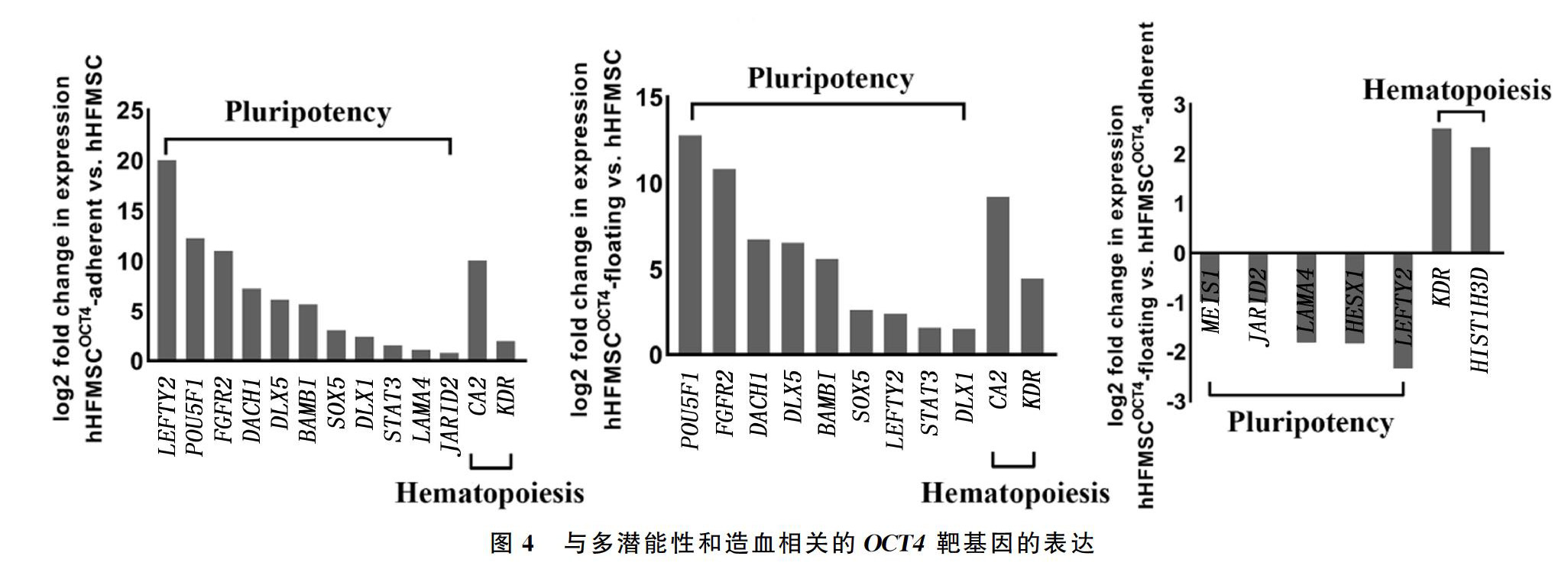
在转导OCT4之后,hHFMSCOCT4-adherent和hHFMSCOCT4-floating中OCT4(POU5F1)表达上调倍数分别为4 859.8和6 958.3倍,两组分别有81和96个OCT4的下游靶基因表达发生改变,其中上调基因分别为48和38个,下调基因分别为33和58个;而hHFMSCOCT4-floating与hHFMSCOCT4-adherent相比,有26个OCT4靶基因表达发生改变,其中上调基因4个,下调基因22个。功能分析表明,与hHFMSC相比,hHFMSCOCT4-adherent 中有SOX5、LEFTY2等11个多潜能性相关靶基因表达上调,而hHFMSCOCT4-floating中有LEFTY2、STAT3等9个多潜能性相关靶基因表达上调;相比hHFMSCOCT4-adherent,hHFMSCOCT4-floating中HESX1、LEFTY2等5个多潜能性相关靶基因表达下调。研究结果表明,OCT4转入 hHFMSC后,对基因转录组进行了重编程,通过上调其他多潜能性基因使细胞获得更多分化潜能。另外,促红细胞生成基因CA2和胚胎造血相关基因KDR在贴壁细胞以及悬浮细胞中表达均上调,说明CA2、KDR基因在OCT4促进造血谱系分化的过程中可能发挥重要的作用。直接与hHFMSCOCT4-adherent进行比较,hHFMSCOCT4-floating中部分多潜能相关基因的表达下调,同时,KDR基因表达继续上调,另一个造血相关靶基因HIST1H3D表达也上调,结果表明hHFMSCOCT4-floating可能丢失一定的多潜能性,并同时获得造血谱系分化倾向(图4)。
3讨论
OCT4是维持干细胞自我更新的重要转录因子,可以与靶基因启动子区结合激活其表达[5-6],包括维持胚胎干细胞自我更新有关基因的表达[7]。在调节细胞多潜能性的基因网络中,OCT4发挥核心调控作用[8-10]。OCT4可结合自身基因及其他多潜能性基因NANOG、SOX2、SALL4等的启动子并调节其转录表达[5-6];另一方面,OCT4与抑制性基因的启动子相结合可抑制其转录激活,而这些抑制性基因的主要功能是促进谱系特异性分化[11-12]。已有研究证实,OCT4表达下调时,与内胚层谱系发育和细胞分化相关的基因转录水平显著升高[13],因此OCT4对于胚胎干细胞多潜能性的维持和成体细胞重编程均具有关键作用[14-15]。此外,最近有研究结果证实,单独转导OCT4即可赋予成纤维细胞向三胚层谱系转分化的“可塑性”(不是完全去分化的多潜能性),即转导OCT4可以上调造血、神经发育及内胚层发育相关基因的表达,在进一步的谱系特异性信号刺激下分别向中胚层(造血祖细胞)和外胚层(神经祖细胞)分化[16-17]。并且,与其他多潜能基因(NANOG或SOX2)相比,只有过表达OCT4才能产生全新的中间状态细胞(非iPSC),并形成造血样克隆(表达CD45)[18],表明OCT4对于体细胞向造血谱系分化具有重要作用。
继转导多个转录因子重编程体细胞使其获得多潜能性后[19-20],近些年研究发现,单独转导OCT4就可重编程体细胞[21]。之前研究中,需要通过慢病毒向体细胞中转导多个重编程基因,其诱导过程复杂,而且导入多因子可能影響基因组稳定性,因此应用具有一定局限性。hHFMSC来源于毛囊,具有来源丰富、获取简单、无免疫原性等应用优点[12],因此可以作为基因工程细胞进行重编程、转分化等研究。本研究通过RNA测序在转录水平检测转导OCT4后转录组的变化,探讨OCT4对hHFMSC分化潜能的影响及其分子机制。
本研究中,单独转导OCT4入hHFMSC后产生贴壁细胞和悬浮细胞两个亚群,与hHFMSC相比,这两种细胞的基因转录本均发生显著变化,筛选出DEGs并进一步进行功能分析显示,上调DEGs显著富集于外胚层和中胚层各器官发育及细胞分化的生物过程中,并且显著富集于调节干细胞多潜能性的信号通路中,说明OCT4重编程的细胞亚群获得了跨谱系、跨胚层分化的多潜能性。直接比较贴壁细胞和悬浮细胞则显示,这两种细胞为相对独立的亚群,具有差异显著的转录组,尽管其差异性不及OCT4转导前后的变化。本研究结果还显示,悬浮细胞亚群中OCT4的上调倍数明显高于贴壁细胞,推测OCT4的较高表达量可能与悬浮细胞的产生有关;另一方面,贴壁细胞经过传代培养后黏附性降低、细胞变圆,而悬浮细胞也可以重新贴壁培养并形成集落[3],说明两种亚型之间能相互转化衍生,其具体机制尚待研究。对OCT4下游靶基因进行分析表明,hHFMSC中OCT4可能通过上调多潜能性相关靶基因LEFTY2、SOX5、FGFR2及STAT3等的表达,使hHFMSC重编程并获得更多的分化潜能,而悬浮细胞中一部分多潜能性靶基因表达量下降,表明该细胞亚群的自我更新与分化的平衡有所倾斜,可能更易于发生转分化。在OCT4的靶基因中,胚胎造血相关基因KDR与红系生成相关基因CA2及造血基因HIST1H3D均表达上调,说明悬浮细胞具有一定的红系分化倾向。因此,在给予造血相关细胞因子刺激后,悬浮细胞最终能够转分化为成熟的红细胞[3]。然而,悬浮细胞与贴壁细胞多潜能性及其他生物学功能的差异仍需进一步探讨,OCT4转导后的细胞具有哪些组织特异性细胞分化能力也需进行相关研究证实。
本研究通过RNA测序方法探讨过表达OCT4后hHFMSC基因转录组的变化及其对分化潜能的影响,寻找OCT4通过调控靶基因表达使体细胞去分化的分子依据,为hHFMSC在基因工程研究中的应用奠定了基础,也为临床应用干细胞进行个体化治疗提供了新的细胞来源。
[参考文献]
[1]ZABIEROWSKI S E, BAUBET V, HIMES B, et al. Direct reprogramming of melanocytes to neural crest stem-like cells by one defined factor[J].? Stem Cells Dayt Ohio, 2011,29(11):1752-1762.
[2]ZHOU J, HOU Y P, ZHANG Z Y, et al. Conversion of human fibroblasts into functional Leydig-like cells by small molecules and a single factor[J].? Biochemical and Biophysical Research Communications, 2019,516(1):1-7.
[3]LIU Z J, LU S J, LU Y, et al. Transdifferentiation of human hair follicle mesenchymal stem cells into red blood cells by OCT4[J].? Stem Cells International, 2015,2015:389628.
[4]WANG Y M, LIU J Y, TAN X H, et al. Induced pluripotent stem cells from human hair follicle mesenchymal stem cells[J].? Stem Cell Rev Rep, 2013,9(4):451-460.
[5]JAENISCH R, YOUNG R. Stem cells, the molecular circuitry of pluripotency and nuclear reprogramming[J].? Cell, 2008,132(4):567-582.
[6]YOUNG R A. Control of the embryonic stem cell state[J].? Cell, 2011,144(6):940-954.
[7]NIWA H. How is pluripotency determined and maintained[J]? Dev Camb Engl, 2007,134(4):635-646.
[8]NICHOLS J, ZEVNIK B, ANASTASSIADIS K, et al. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4[J].? Cell, 1998,95(3):379-391.
[9]SHI G L, JIN Y. Role of Oct4 in maintaining and regaining stem cell pluripotency[J].? Stem Cell Res Ther, 2010,1(5):39.
[10]VAN DEN BERG D L C, SNOEK T, MULLIN N P, et al. An Oct4-centered protein interaction network in embryonic stem cells[J].? Cell Stem Cell, 2010,6(4):369-381.
[11]BOYER L A, LEE T I, COLE M F, et al. Core transcriptio-nal regulatory circuitry in human embryonic stem cells[J].? Cell, 2005,122(6):947-956.
[12]LOH Y, WU Q, CHEW J L, et al. The Oct4 and Nanog transcription network regulates pluripotency in mouse embryonic stem cells[J].? Nature Genetics, 2006,38(4):431-440.
[13]ZAFARANA G, AVERY S R, AVERY K, et al. Specific knockdown of OCT4 in human embryonic stem cells by indu-cible short hairpin RNA interference[J].? Stem Cells Dayt Ohio, 2009,27(4):776-782.
[14]LE BIN G C, MUNOZ-DESCALZO S, KUROWSKI A, et al. Oct4 is required for lineage priming in the developing inner cell mass of the mouse blastocyst[J].? Development, 2014,141(5):1001-1010.
[15]WERNIG M, MEISSNER A, FOREMAN R, et al. In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state[J].? Nature, 2007,448(7151):318-324.
[16]MITCHELL R R, SZABO E, BENOIT Y D, et al. Activation of neural cell fate programs toward direct conversion of adult human fibroblasts into tri-potent neural progenitors using OCT-4[J].? Stem Cells Dev, 2014,23(16):1937-1946.
[17]RING K L, TONG L M, BALESTRA M E, et al. Direct reprogramming of mouse and human fibroblasts into multipotent neural stem cells with a single factor[J].? Cell Stem Cell, 2012,11(1):100-109.
[18]SZABO E, RAMPALLI S, RISUEO R M, et al. Direct conversion of human fibroblasts to multilineage blood progenitors[J].? Nature, 2010,468(7323):521-526.
[19]TAKAHASHI K, YAMANAKA S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors[J].? Cell, 2006,126(4):663-676.
[20]YU J, VODYANIK M A, SMUGA-OTTO K, et al. Induced pluripotent stem cell lines derived from human somatic cells[J].? Science (New York, NY), 2007,318(5858):1917-1920.
[21]MITCHELL R, SZABO E, SHAPOVALOVA Z, et al. Molecular evidence for OCT4-induced plasticity in adult human fibroblasts required for direct cell fate conversion to lineage specific progenitors[J].? Stem Cells, 2014,32(8):2178-2187.
(本文編辑 马伟平)
- 奧尔夫音乐教学法形成的方法论原则
- 琵琶在晋南戏曲音乐伴奏中的现状调查
- 《玩家》
- 我看英文原版音乐剧《芝加哥》
- 小剧种与大角色
- 一台锣鼓半台戏
- 校园话剧《艺海丰碑》艺术特色之论析
- 精神分析学视域下曹禺《原野》中仇虎心理演变
- 襄阳大越调皮影戏艺人李有生口述史
- 印第安文化和谐之解读
- 动画电影《寻梦环游记》的创作理念研读
- 央视春晚主、分会场直播模式的历史沿革及对井冈山春晚分会场的启示
- 我国影视产品走向国际市场的现状与障碍因素分析
- 香港电视剧的创新之路
- 探究微电影时空设计与后现代编剧创构
- 从艺术学角度探讨影视批评的意义与价值
- 以电影《聚焦》背后真实事件为例谈媒体的舆论监督作用
- 《我不是药神》的存在主义之思
- 论《红高粱》中象征手法的运用
- 浅谈影视表演学生的现状及其发展方向
- 成长中的“他者”建构与“自我”实现
- 何谓纪录片
- 《明星大侦探》:网络综艺节目的战略分析
- 性别政治视域下女性意识表达的理性审视
- “为有牺牲多壮志《右玉和她的县委书记们》”
- make sb's acquaintance
- make sb's acquaintance / make the acquaintance of sb
- make sb sad/unhappy
- make sb's day
- make sb sth
- make sb/sth into sb/sth
- make sb/sth out
- make sb think of
- make sb unsure/uncertain
- make sb wait
- make sense
- make-sense
- make sense of
- makes-for
- makeshift
- makeshiftness
- makeshifts
- makeshifty
- make-short-work-of
- make somebody out
- make somebody/somethingout
- make somethingout
- make somethingup
- makes out
- makes-over
- 超级模特
- 超级癌症
- 超级稻
- 超级管弦乐团
- 超纲
- 超纲要
- 超线性成分
- 超经济型酒店
- 超绝
- 超绝不凡
- 超绝出众
- 超绝古今
- 超绝尘寰
- 超绝迈越常流
- 超绝,妙极
- 超绝,超出
- 超编
- 超群
- 超群出众
- 超群出众的人或物
- 超群出众的人才
- 超群出众的美丽容貌
- 超群出众的风度气质
- 超群出众,不同一般
- 超群出众,位居第一