宋立民 李明泽 李仰真 马林 吴会民 刘义 李楠
摘要 為评价遗传背景、体重、黑化、运输应激等对牙鲆(Paralichthys olivaceus)血清免疫学指标的影响,以引进的“鲆优2号”抗病牙鲆和普通牙鲆为试验材料,通过肉眼区分无眼侧有无黑化,在不同生长阶段采集样本,采样前有无施行包装运输等手段,将研究对象分为7个试验组,分别测定溶菌酶(LYS)活力、酸性磷酸酶(ACP)活力、超氧化物歧化酶(SOD)活力、免疫球蛋白M(IgM)含量、补体3(C3)和补体4(C4)含量等6种免疫学指标。结果表明:试验组1各项检测指标均低于试验组2,但差异均不显著(P>0.05);试验组4各项检测指标均低于试验组7,其中LYS、ACP、SOD活力及IgM含量差异极显著(P<0.01),C4含量差异显著(P<0.05),C3含量差异不显著(P>0.05);试验组1各检测指标均低于试验组7,且差异超级显著(P<0.001);试验组2各检测指标均小于试验组4,SOD活力及IgM、C3含量差异超级显著(P<0.001),LYS活力差异极显著(P<0.01),C4含量差异显著(P<0.05),ACP活力无显著差异;试验组3与试验组4各指标差异均不显著(P>0.05);比较试验组5、试验组6与试验组7的检测结果发现,只有试验组5中C3含量极显著低于试验组6(P<0.01),其余指标差异均不显著。通过一定的育种手段可使养殖对象在某一个或多个方面获得可遗传的免疫优势;随着牙鲆个体的生长,体重增加,抗病力逐渐增强;牙鲆的抗病能力与黑化与否的相关性未达到显著水平;对于同一遗传背景,经历运输(未产生休克)与未经历运输样本鱼的6种免疫学指标差异不显著。
关键词 牙鲆;遗传背景;体重;黑化;运输应激;免疫学指标
中图分类号 S917.4文献标识码 A文章编号 0517-6611(2020)22-0094-05
doi:10.3969/j.issn.0517-6611.2020.22.025
Comparison of Serum Immune Indices of Paralichthys olivaceus under Different Conditions
SONG Li-min1, LI Ming-ze1, LI Yang-zhen2 et al
(1. Tianjin Fishery Research Institute, Tianjin 300221;2. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences (CAFS), Qingdao,Shandong? 266071)
Abstract In order to evaluate the influence of genetic background, body weight, hypermelanosis and transportation stress on the serum immune indices of Paralichthys olivaceus, the subjects were divided into 7 experimental groups, according to blind-side with hypermelanosis or not, different growth stages, and sampling with transportation stress or not, from the resistant flounder and common flounder which were used as experimental materials. Lysozyme (LYS) activity, acid phosphatase (ACP) activity, superoxide dismutase (SOD) activity, immunoglobulin M (IgM) content, complement C3 content, C4 content were measured and compared separately of the above groups. The results indicated that the test items of experimental group 1 were less than those of group 2, but the difference was not significant (P>0.05).The test items of experimental group 4 were lower than those of group 7, and there were highly significant differences (P<0.01) in LYS activity, ACP activity, SOD activity and IgM content, while the difference in C4 content was significant (P<0.05), there was no significant difference (P>0.05) in C3 content.The test items of experimental group 1 were lower than those of group 7, and the difference were super significant (P<0.001).The test items of experimental group 2 were lower than those of group 4, and there were super significant differences (P<0.001) in SOD activity, IgM content and C3 content, while highly significant difference (P<0.01) in LYS activity, significant difference (P<0.05) in C4 content, no significant difference (P>0.05) in ACP activity.There were no significant difference (P>0.05) in 6 immune indices between experimental group 3 and group 4.Only C3 content of group 5 was highly significant (P<0.01) lower than that of group 6, by comparing the test items of experimental group 5, group 6 and group 7. The results showed that heritable immune advantage in one or more aspects could be obtained by cultured animals, according to certain breeding methods.The disease resistance was gradually increased with the growth of Paralichthys olivaceus individuals.There was no significant correlation between immune indices and blind-side with hypermelanosis or not of Paralichthys olivaceus.The same result as above was obtained about sampling with transportation stress (which was not an approaching death shock) or not in this experiment.
Key words Paralichthys olivaceus;Genetic background;Body weight;Hypermelanosis;Transportation stress;Immune indices
基金项目 天津市农业发展服务中心青年科技创新项目(ZXKJ201908,ZXKJ201911)。
作者简介 宋立民(1982—),男,河北秦皇岛人,工程师,硕士,从事水产养殖与育种研究。*通信作者,工程师,硕士,从事水产养殖研究。
收稿日期 2020-05-31
牙鲆(Paralichthys olivaceus)属于鲽形目鲽亚目牙鲆科牙鲆属,以黄渤海区分布较多,其主要捕食底栖非经济鱼类及甲壳类,肉质细嫩鲜美[1],是我国重要的海水养殖品种之一。鉴于牙鲆养殖过程中,周年可发生腹水病害,研究表明其主要致病菌之一是迟缓爱德华氏菌(Edwardsiella tarda)[2-4],陈松林团队通过多年努力培育出抗迟缓爱德华氏菌病能力强、生长较快的牙鲆新品种——“鲆优2号”,为减少养殖牙鲆病害发生、发展无抗养殖提供了新思路[5]。笔者所在课题组于2018年引进“鲆优2号”牙鲆抗病苗种,试养期间采集了新品种牙鲆和普通牙鲆的血清样本,用于测定其溶菌酶活力(LYS)、酸性磷酸酶活力(ACP)、超氧化物歧化酶活力(SOD)、免疫球蛋白M含量(IgM)、补体3(C3)和补体4(C4)含量等重要免疫学指标,依据牙鲆不同遗传背景、不同体重、无眼侧有无黑化、采样前有无运输应激等分成不同试验组,并对测定的免疫学指标进行比较,以期为“鲆优2号”牙鲆新品种的抗病原理研究提供免疫学依据,同时为进一步研究牙鲆免疫学指标的影响因素积累数据。
1 材料与方法
1.1 试验鱼来源及饲养条件
“鲆优2号”牙鲆新品种购于海阳市黄海水产有限公司,普通牙鲆购于秦皇岛启民水产养殖有限公司,二者饲养条件相同。试验地位于天津市民峰水产有限公司,使用饲料为莱州鸿颐水产有限公司生产的“海童”牌鲆鲽类配合饲料,水泥池面积40 m2,苗种规格为200 g左右时放养密度为32尾/m2,苗种规格为600 g左右时放养密度为17尾/m2,养殖方式为流水养殖,水温18.0~21.5 ℃,溶解氧含量6.14~8.02 mg/L,pH 8.0~8.2,盐度26‰。
1.2 试验分组与样本采集
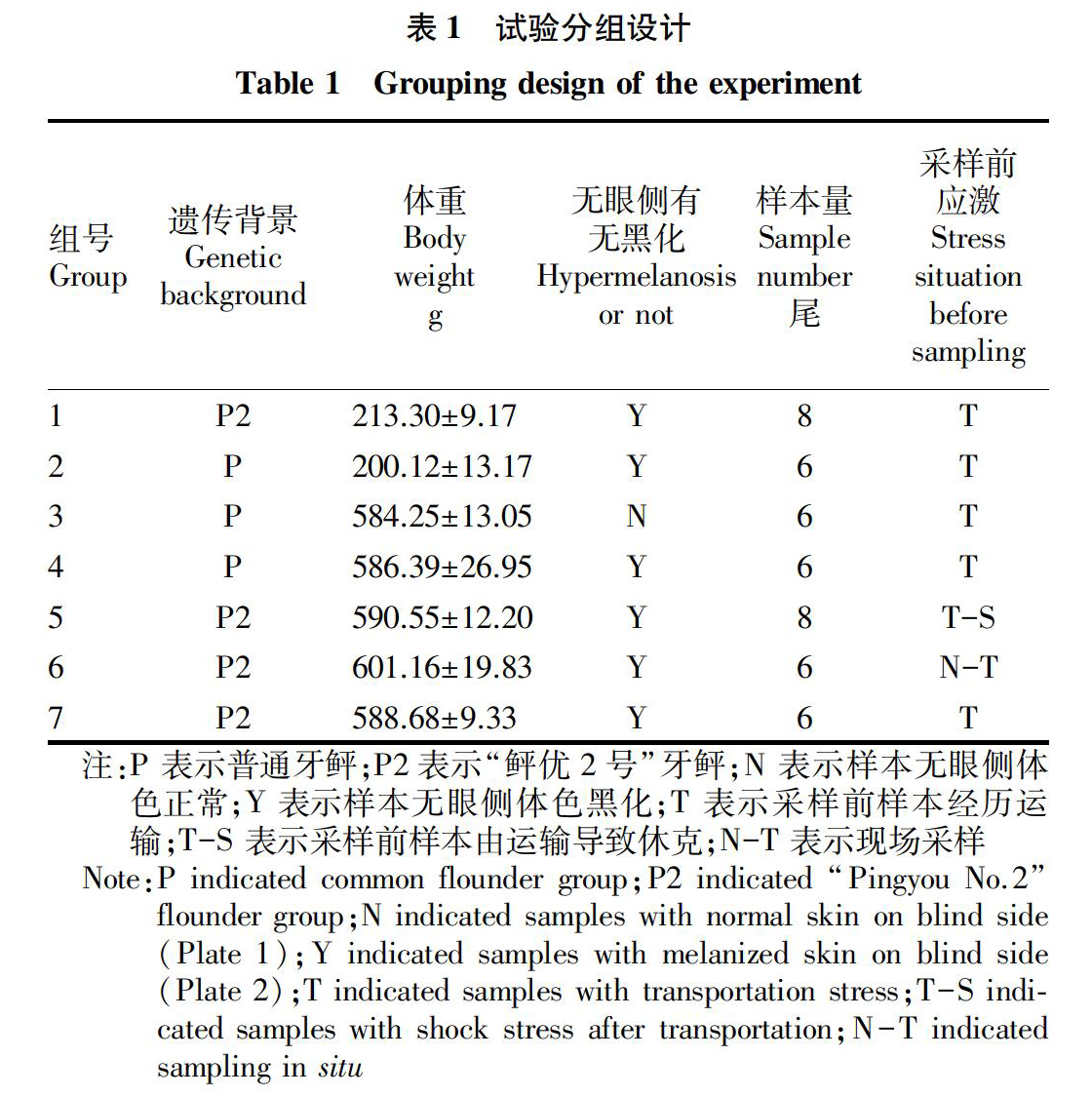
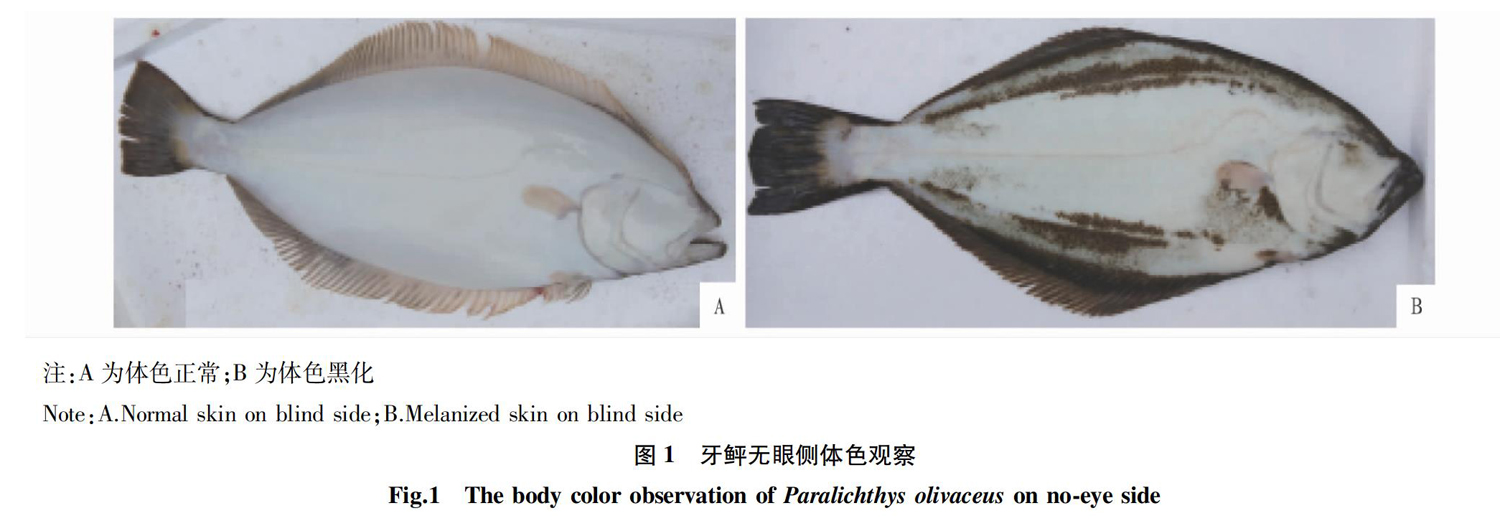
依据遗传背景,分为普通牙鲆和选育的“鲆优2号”牙鲆新品种两大群体;依据体重规格,可分为200和600 g两个生长阶段。依据无眼侧有无黑化(图1),从普通牙鲆苗群体中分别采集黑化样本和正常样本;另外依据采样前有无进行运输应急,从牙鲆新品种群体中采集应激样本和未应激样本,具体情况如表1所示。
采样前进行打包运输操作,参照米红波等[6]的方法并根据实际条件进行调整。运输包装采用透明PE平口袋(长90 cm、宽60 cm),运输用水采用曝气井水,pH 8.3,水温21.5 ℃,盐度26‰,溶解氧含量8.02 mg/L。包装袋内注入1/3的水,每袋放入2尾牙鲆,充气,封口。历时80 min运至实验室。表1中试验组5样本规格较大,分4次打包,历时110 min运至实验室,其中6尾无呼吸动作、产生休克,另2尾呼吸动作虚弱,将休克鱼送至实验室,立即采集血样;试验组6样本没有经历运输应急,试验鱼从养殖车间现场捞取,直接进行血液采集。
1.3 血清样本的制备和部分免疫学指标的测定
血清样本采集参考姜宏波[7]方法,并做适当改动。具体方法如下:试验鱼禁食24 h后,使用200~400 μL/L乙二醇苯醚麻醉,用2 mL注射器于尾部静脉迅速抽取1.0~1.5 mL血液,缓慢注入2 mL无菌离心管中,于4 ℃冰箱中静置2 h,以3 000 r/min 转速离心15 min,分离获得上清液(即血清);用0.2 mL无菌离心管分装密封,-80 ℃下冷冻待测。保存过程中若出现沉淀,则必须再次离心,重新收集上清液。试验所需试剂盒购于上海纪宁实业有限公司,使用Rayto RT-6100酶标分析仪进行部分免疫学指标检测。检测指标包括溶菌酶(LYS)活力、酸性磷酸酶(ACP)活力、超氧化物歧化酶(SOD)活力、免疫球蛋白M(IgM)含量、补体蛋白3(C3)含量、补体蛋白4(C4)含量。
1.4 数据处理
试验数据均以平均值±标准差(Mean ± SD)表示。使用SPSS 13.0统计软件对试验数据进行统计分析,P<0.05表示差异显著,P<0.01表示差异极显著,P<0.001表示差异超级显著,使用Excel 2010软件绘制相关图表。
2 结果与分析
2.1 不同遗传背景对牙鲆血清免疫学指标的影响
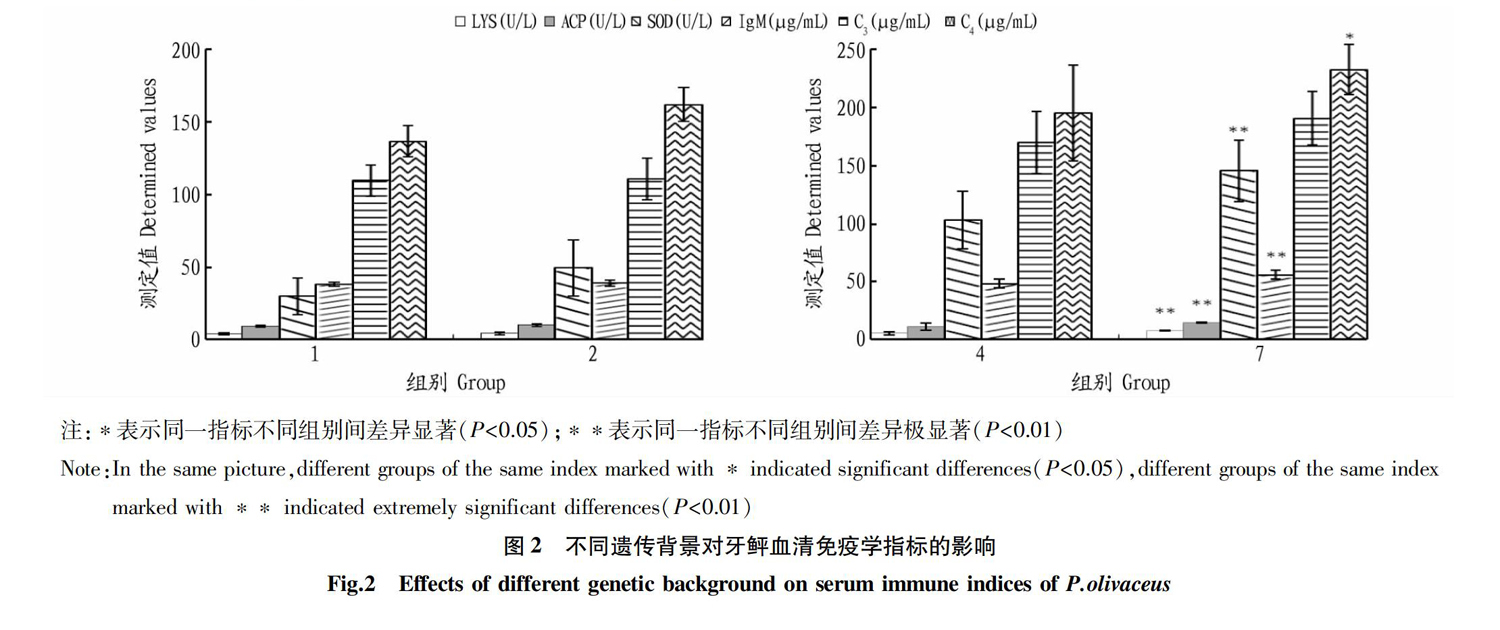
从图2可以看出,试验组1各项检测指标均低于试验组2,但差异均不显著(P>0.05)。试验组4各项检测指标均低于试验组7,其中LYS活力、ACP活力、SOD活力、IgM含量差异极显著(P<0.01);C4含量差异显著(P<0.05);C3含量差异不显著(P>0.05)。试验组1、试验组4是从秦皇岛启民水产养殖有限公司购得的普通牙鲆,试验组2、试验组7是从海阳市黄海水产有限公司购得的选育新品种“鲆优2号”,试验组1、4与试验组2、7遗传背景不同,检测结果表明经过选育的“鲆优2号”牙鲆上述血清免疫学指标比普通牙鲆更有优势,并随着个体的生长发育,新品种免疫学优势呈增强趋势。
2.2 不同体重对牙鲆血清免疫学指标的影响
从图3可以看出,试验组1各项检测指标均低于试验组7,且差异超级显著(P<0.001);试验组2各项检测指标均低于试验组4,SOD活力、IgM含量、C3含量差异超级显著(P<0.001),LYS活力差异极显著(P<0.01),C4含量差异显著(P<0.05),ACP活力无显著差异(P>0.05)。试验组1牙鲆平均体重(213.30±9.17)g,试验组2平均体重(200.12±13.17)g,二者差异不显著(P>0.05);试验组4牙鲆平均体重为(586.39±2695)g,试验组7平均体重为(588.68±9.33)g,二者差异不显著(P>0.05);试验组1、7与试验组2、4牙鲆的生长阶段不同,依照试验总体结果,随着牙鲆个体的生长,其血清免疫学指標产生变化,抗病力逐渐增强,这种趋势在“鲆优2号”牙鲆群体中表现尤为明显。
2.3 无眼侧黑化与正常牙鲆血清免疫学指标比较
从图4可以看出,试验组3、4试验鱼均为购自秦皇岛启民水产养殖有限公司的普通牙鲆,试养环境相同,无眼侧体色不同,各检测指标差异均不显著(P>0.05),说明牙鲆无眼侧黑化与否与所测指标在血清中的水平无显著相关。
2.4 不同运输应激对牙鲆免疫学指标的影响
从图5可以看出,试验组5、6与试验组7牙鲆体重差异不显著,采样前样本鱼经历操作方式不同,各检测指标中,只有试验组5中C3含量极显著低于试验组6(P<0.01),其余值差异均不显著。样本鱼在短暂运输过程中没有对检测指标产生明显影响,但如果样本鱼受到一定程度的应激(比如发生休克),则会对某些指标产生显著影响,其中C3含量在6种检测指标中表现更为敏感。
3 讨论
3.1 選择6种指标初步评价免疫力的原因
LYS在抵抗外来病原入侵中起着重要的作用[8]。LYS能破坏细胞壁中的肽聚糖,使细菌细胞崩解[9];此外,它还能激活补体旁路和吞噬细胞活性,参与机体非特异性免疫防御[10]。LYS 是非常重要的免疫因子,其活性高低反映水生生物非特异性免疫水平的高低,因此有很多研究都采用LYS作为重要的免疫学指标[11-13]。ACP广泛分布于生物界[13],是巨噬细胞溶酶体的标志酶之一,参与动物机体的免疫活动[14-15]。SOD广泛分布于机体血液和各组织器官中,是生物体中最先对活性氧自由基作出反应的抗氧化酶[16]。免疫球蛋白是鱼类特异性体液免疫应答的主要介质[17]。目前鱼类是最早产生免疫球蛋白的脊椎动物,可能仅有免疫球蛋白IgM[14]。IgM含量通常被认为是评价鱼体免疫应答反应的重要指标[9]。鱼类的补体系统通过吞噬细胞协助作用来溶解外来细胞和外来生物体[18],同时参与特异性和非特异性免疫[19],其中C3和C4在补体系统中担当着重要角色[20]。因此,该试验选择上述6种指标来评估牙鲆免疫水平。
3.2 不同遗传背景对牙鲆免疫力的影响
姜宏波等[21]认为血液中的一些指标是反映机体健康状况及抗病力的重要参数,对克隆牙鲆与普通牙鲆血液生理生化指标的研究表明遗传因素是影响这些指标的重要方面。田岳强等[22]对大菱鲆(Scophthalmus maximus)选育家系与普通家系LYS、IgM等7种非特异免疫因子在肝脏、脾脏与头肾中的表达量研究表明攻毒前后6 个选育家系免疫因子的表达量均高于普通组。刘堃等[23]研究表明抗病草鱼(Ctenopharyngodon idellus)血液中免疫球蛋白IgM转录本丰度、LYS含量等部分免疫学生化指标显著高于普通草鱼(P<0.05)。杨启莲等[24]研究表明选育的“闽优1 号”大黄鱼(Larimichthys crocea)血清LYS活力和血细胞SOD活力均显著高于对照组(P<0.05),而血清中的免疫球蛋白含量低于普通养殖群体。该试验在生理水平上同样证明经过相同阶段试养,人工选育的“鲆优2号”牙鲆新品种比普通牙鲆更具免疫优势,只是不同生长阶段优势差异程度不同,而且各评价指标随着生长发育的提升程度亦不同,如试验中补体C3含量的变化程度相对其他5种指标较小,可能是由于其自身生理机制的特点以及定向选育操作的影响。综上所述,通过选择育种等手段可以使养殖对象在某一个或多个方面获得可遗传的免疫优势,但不同指标在不同种类、机体不同部位中的表达结果存在差异,这需要对免疫应答机理进行更深入研究。
3.3 不同体重对牙鲆免疫力的影响
李泽宇[25]在建立牙鲆抗迟缓爱德华氏菌病家系的研究中发现牙鲆鱼的全长、体重等生长性状与抗病力呈极显著正相关。对哺乳动物猪的研究发现,仔猪体重越大,免疫力和抗病力越强[26],且相同生长期大个体的抗病力要强于小个体。该试验养殖过程中,随着牙鲆个体的生长,体重增加,抗病力逐渐增强,与前述研究结果类似,然而也存在与该试验不同的研究结果。侯吉伦等[27]在牙鲆抗淋巴囊肿病家系的选育过程中发现牙鲆淋巴囊肿抗病和患病个体间的体重和体长差异均不显著,这可能与抗病性能评判标准不同有关。该试验是以血清中几种免疫相关酶为评价指标,间接评估试验组鱼的抗病力,而李泽宇[25]是以试验牙鲆人工感染迟缓爱德华氏菌后的成活率,侯吉伦等[27]是以在患病环境中饲养、出现淋巴囊肿症状为标准进行评价,同时不同致病菌造成机体免疫应答机制产生差异也是可能原因之一。穆秀琼[28]在对吉富罗非鱼的养殖试验表明,7周后各家系的总超氧化物歧化酶活力、LYS活力分别与体重增长呈显著正相关,即体重增长较快的家系其总超氧化物歧化酶水平也较高;试验14周和21周后却表现为体重的增长分别与SOD活力、LYS活力的变化没有显著相关性[28]。这可能与选取的试验对象种类、生长阶段和养殖方式不同有关。依照该试验结果,“鲆优2号”牙鲆群体中体重增加对血清免疫学指标产生的影响比普通群体更为明显,这可能与“鲆优2号”牙鲆经过选育个体差异较小,所获得抗病力随着体重的增长而逐渐增强有关。
3.4 无眼侧有无黑化对牙鲆免疫力的影响
“黑化”指的是无眼侧出现黑斑,即黑色素细胞增加。养殖环境、密度、营养等都能够对牙鲆的体色产生影响[29-30],亦能够直接或间接影响鱼类免疫力[31-33]。相关研究表明,金黄壳色马氏珠母贝(Pinctada fuctada martensii)群体与养殖群体血清免疫酶活力存在差异[34],3种颜色刺参(Apostichopus japonicus Selenka)在不同光照强度下的免疫酶活性存在差异[35],另外鉴于该试验引进的“鲆优2号”幼苗无眼侧黑化情况较为普遍,对照普通苗中存在一定比例的黑化和正常苗;对不同体色牙鲆之间的免疫学指标进行了比较,结果表明牙鲆的抗病力与无眼侧黑化与否的关联性不显著。
3.5 运输应激对牙鲆免疫力的影响
在活鱼运输过程中水体污染、振动、密度过高、温度变化、运输时间过长等因素均会不同程度引发鱼体应激,进而影响机体功能[36]。宋凯等[37]对牙鲆的研究表明运输将打破鱼体的内分泌平衡,使其机体内产生氧化应激。该试验结果表明,相同背景、经历运输(未产生休克)与未经历运输样本鱼的6种免疫学指标差异不显著,究其原因可能是由于机体受到应激胁迫后,首先进行一级、二级反应,严重时才导致三级反应,即免疫系统受到抑制等[38-39],上述6种免疫学指标在机体内参与应激反应的时间相对滞后。
4 结论
综上所述,通过一定的育种手段可使新品种牙鲆获得一定的免疫优势;随着牙鲆个体的生长,体重增加,抗病力显著增强;牙鲆的抗病性能与黑化与否的关联性不显著;经过运输(未产生休克)与未经过运输样本鱼的6种免疫学指标差异不显著。该试验结果表明定向育种操作、体重、休克均可对牙鲆某种或某几种免疫学指标产生影响,无眼侧有无黑化、适度的运输操作对该试验中牙鲆6种免疫学指标的影响不显著。
参考文献
[1] 宋立民,于清海,袁立来,等.回捕牙鲆生长和摄食状况调查研究[J].天津农业科学,2019,25(5):33-38.
[2] 尚琨,曲凌云,王玉芬,等.患腹水病牙鲆病原菌分离、鉴定及病原菌的特性[J].水产学报,2020,44(2):266-275.
[3] 朱壮春,史相国,张淑杰,等.牙鲆腹水病病原研究[J].水产科学,2006,25(7):325-329.
[4] 周丽,宫庆礼,俞开康.牙鲆的疾病[J].青岛海洋大学学报,1997,27(2):173-180.
[5] 陈松林.鱼类基因组学及基因组育种技术[M].北京:科学出版社,2017.
[6] 米红波,侯晓荣,茅林春.鱼虾类保活运输的研究与应用进展[J].食品科学,2013,34(13):365-369.
[7] 姜宏波.双单倍体牙鲆纯合不育个体的蛋白质组和转录组学研究[D].哈尔滨:东北农业大学,2015.
[8] 郑清梅,吴锐全,叶星.水生动物溶菌酶的研究进展[J].上海水产大学学报,2006,15(4):483-487.
[9] 谢明媚,彭士明,张晨捷,等.急性温度胁迫对银鲳幼鱼抗氧化和免疫指标的影响[J].海洋渔业,2015,37(6):541-549.
[10] ZHU L Y,NIE L,ZHU G,et al.Advances in research of fish immune-relevant genes:A comparative overview of innate and adaptive immunity in teleosts[J].Developmental & comparative ummunology,2013,39(1/2):39-62.
[11] 王树芹,周洪琪.壳聚糖对异育银鲫溶菌酶和白细胞吞噬活性的影响[J].上海水产大学学报,2004,13(2):121-125.
[12] 徐大伦,黄晓春,欧昌荣,等.浒苔多糖对华贵栉孔扇贝血淋巴中SOD酶和溶菌酶活性的影响[J].水产科学,2006,25(2):72-74.
[13] 常杰.对虾和刺参敏感免疫学指标的筛选和评价[D].青岛:中国海洋大学,2010.
[14] 陈启明.精氨酸对黄颡鱼生长、免疫及肠道健康的影响[D].广州:华南农业大学,2016.
[15] SONG S K,BECK B R,KIM D,et al.Prebiotics as immunostimulants in aquaculture:A review[J].Fish & shellfish immunology,2014,40(1):40-48.
[16] 孙俊霄,韩广坤,刘娅,等.杂交黄颡鱼与普通黄颡鱼幼鱼生长性能及耐低氧能力的比较[J].水生生物学报,2019,43(6):1271-1279.
[17] 张立颖,赵萌.鱼类免疫球蛋白的研究进展[J].水产科学,2009,28(11):701-705.
[18] HOLLAND M C H,LAMBRIS J D.The complement system in teleosts[J].Fish & shellfish immunology,2002,12(5):399-420.
[19] 王俊相,李玉萍,孔令富,等.魚类免疫系统的研究进展[J].四川畜牧兽医,2010,37(7):29-31.
[20] KURODA N,NARUSE K,SHIMA A,et al.Molecular cloning and linkage analysis of complement C3 and C4 genes of the Japanese medaka fish[J].Immunogenetics,2000,51(2):117-128.
[21] 姜宏波,王桂兴,刘海金,等.克隆牙鲆和普通牙鲆血液生理生化指标的比较[J].中国水产科学,2014,21(2):260-265.
[22] 田岳强,郭建丽,黄智慧,等.大菱鲆家系选育二代7种免疫因子的分析[J].海洋科学,2016,40(9):9-17.
[23] 刘堃,陈剑兴,陈胜峰,等.雌核发育抗病草鱼与普通草鱼的免疫学指标比较[J].生命科学研究,2015,19(3):213-217.
[24] 杨启莲,姚翠鸾,王志勇.大黄鱼选育群体与普通养殖群体部分免疫指标的比较[J].海洋科学,2012,36(10):48-53.
- 肺炎疫苗联合流感疫苗在老年人群中的作用
- 外科快速康复理念在脑内血肿患者术后临床护理中的应用观察
- 分析对重症患者气管插管进行精心护理的必要性
- 预防手术患者术中低体温的护理措施
- 对优质护理管理应用在腹腔镜成人疝修补术中的应用效果进行分析
- ICU重症监护室建立人工气道患者有效护理方法研究
- 采血室针刺伤的原因分析及解决方式分析
- 综合护理干预和常规护理在子宫肌瘤手术中的护理效果对比观察
- 护理风险干预预防重症患者压疮的效果
- 集束化护理对大便失禁重症患者失禁性皮炎发生的影响分析
- 系统健康教育对枫桥街道高血压患者知信行的干预效果研究
- 引导式教育理念在脑瘫儿童康复护理中的实施效果探讨
- 康复运动联合肌肉放松训练在慢性心衰患者中的应用
- 延伸护理服务对主动脉夹层术后患者血压控制的影响
- 快速康复外科护理在高龄髋关节置换患者围手术期中的应用
- 临床护理路径应用于体检者13C尿素呼气试验检查中的效果探究与分析
- 品管圈活动对降低神经内科住院患者跌倒坠床发生率的效果评估
- 早期康复护理治疗腰椎间盘突出症术后的效果分析
- 优质护理在妇产科护理中的应用探讨
- 早期康复护理治疗缺血性脑卒中的效果分析
- 个性化双心护理在冠心病心绞痛患者护理中的研究
- 两种口腔护理溶液应用于口腔护理的效果比较
- 穴位配合乳房按摩对缓解乳房胀痛促进母乳喂养的护理效果观察
- 一对一责任制助产护理对初产妇分娩质量以及护理满意度的影响分析
- 院前急救护理对腹部外伤患者的抢救效果分析
- couscouses
- cousin
- cousinage's
- cousiness
- cousinhood
- cousinhoods
- cousin-hoods
- cousinliness
- cousinly
- cousinry
- cousins
- cousinship
- cousinships
- covalent
- covalents
- cove
- coved
- covelet
- covenant
- covenantal
- covenanted
- covenanting
- covenants
- coventry
- cover
- 主题
- 主题主语
- 主题义界
- 主题促销
- 主题公园
- 主题分析
- 主题变奏式结构
- 主题句
- 主题在国语中的功能研究——迈向言谈分析的第一步
- 主题思想
- 主题性人物
- 主题推进
- 主题歌
- 主题派对
- 主题的多义性
- 主题的开拓
- 主题的提炼
- 主题的揭示
- 主题的深化
- 主题的表现
- 主题结构
- 主题表现式议论
- 主题词
- 主题词群
- 主题语