程晓明 程婧晔 胡文静 蒋正宁


摘要:为了合理利用小麦种质资源,给小麦新品种选育提供材料,收集了国内七大小麦生态区23份品种为试验材料,对茎蘖数、叶绿素含量、苗期株高、拔节期、抽穗期、开花期、成熟期、千粒质量、成穗数、穗粒数10个农艺性状进行主成分及聚类分析。主成分分析表明,从10个农艺性状中可提取拔节期生长因子、开花期生长因子、成穗数产量因子、抽穗期生长因子、千粒质量产量因子共5个主成分,占其总信息量的91.2%。利用这5个主成分因子为综合指标进行系统聚类,在遗传距离为26.64的水平上将23个品种划分成4个类群,即春性弱春性类、冬性半冬性类、强春性类、强冬性类,春化生育特性相近的大部分被分在1个类群,同时表明主成分中以拔节期、抽穗期、开花期构成生长因子,千粒质量、成穗数构成产量因子,较能真实地表现春化生育特性,尤其以拔节期为主效应,至抽穗期、开花期等快速生长阶段表现更具反映春化发育特性的本质。
关键词:小麦;主成分分析;聚类分析;春化特性;生长因子
中图分类号: S512.103文献标志码: A
文章编号:1002-1302(2019)08-0064-04
种质资源的分类是小麦育种研究的重要来源和基础,合理利用小麦种质资源是育种材料的选配及综合评价的前提,北纬33度线地区育种研究,即黄淮小麦生态区南片和长江下游生态地区,北接河南东南部,西部与安徽接壤,地处亚热带向暖温带过渡地区,地域广跨度大,生态条件复杂,土壤类型多样化。北纬33度线地区是我国最重要的小麦粮食产区之一,该地区小麦引种对于选育改良小麦新品种提高粮食产量、保障国家粮食安全具有积极且重要的意义。
以主成分分析为主的聚类分析方法是一种多元统计分析方法,是将原来众多有一定相关性的单个指标组合成1组新的互相独立的主成分单元,在很少损失原有信息的前提下,通过降维将原来的多指标转换为数量较少的彼此独立的新指标,揭示变量间的关系。自20世纪60年代起,应用于油菜的分类研究应用越来越广泛[1-6],Matey等应用聚类分析研究了印度4个自交亲和型油菜的遗传差异[7];蔡健等用系统聚类方法分析了20个皖北主栽小麦品种的遗传多样性[8];张桂英等对山西关中地区92个大田生产小麦品质性状的因子进行了聚类分析[9];刘二明等用系统聚类方法分析了21个水稻品种的抗瘟性[10],实践证明合理利用种质资源、客观评价选配亲本材料,对提高育种效率具有重要的实践意义。20世纪以来,农艺性状鉴定逐渐成为一种广泛应用的方法,即使在分子标记快速发展的今天,充分把握和科学评价种质材料的农艺性状的真实表现并进行分类,对于种质资源的合理利用仍具有重要意义[11]。目前对于小麦种质资源,以小麦生育特性农艺性状类型为基础的春冬性的分类较多,大多集中在一些地方品种,或某些地区的大面积主栽品种,而对于全国大面积范围春冬性差异显著且生育特性类型丰富的各小麦生态区主栽品种的研究较少。
為了进一步丰富北纬33度线地区小麦育种引进种质资源的类型,拓宽小麦育种遗传改良的基础范围,提高和优化选配杂交育种亲本材料的效率及对引进种质资源进行遗传多样性综合评价,本试验收集了我国黑龙江、河北、河南、山东、江苏、山西、陕西、甘肃、宁夏、新疆、四川、云南和西藏等13个省份七大生态区大面积小麦主栽品种23份材料,运用主成分分析及聚类分析方法综合生育特性农艺性状等特点进行春冬性分类研究,旨在为北纬33度线地区育种改良研究决策提供理论依据及数据支持。
1材料与方法
1.1供试材料
供试材料来自全国七大小麦生态区科研院所提供的23份小麦品种,分别是东北春麦区育成品种龙麦33,黄淮冬麦区育成品种石4185、衡观35、郑7698、郑9023、周麦22、西农979、济麦22和烟农19,黄淮麦区南片育成品种小偃22、晋麦47,长江中下游冬麦区育成品种扬麦16、宁麦13、扬麦25,西北春播麦区育成品种兰天26、宁春4号、宁春50号,新疆冬春播麦区育成品种新冬22、新春29,西南麦区育成品种川麦104、绵麦367和云麦53,青藏春冬(播)麦区育成品种肥麦(表1)。
试验前对各品种籽粒进行发芽率试验,试验检验设为2个重复,根据检验数据结果以基本苗225万株/hm2的标准计算确定各品种籽粒播种数量。
1.2试验设计
试验于2016—2107年在江苏里下河地区农业科学研究所试验基地进行,试验点位于长江与京杭大运河交汇处(119°26′E、32°24′N), 具有典型的北亚热带季风气候,温暖湿润无霜期为210~280 d,日平均气温≥10 ℃,积温达4 500~5 600 ℃,年降水量为1 000~1 100 mm。试验田土质为沙壤土,土地平坦。试验于适播期2016年11月5日播种,人工开沟点播。供试每个品种播种3行,行长133 cm,行距23 cm,试验种植密度为基本苗225万株/hm2,重复2次。齐苗后每个品种重复的均定苗1个行长100 cm,均去2侧行头,用于测定成熟期产量构成因素。
1.3田间管理及测定项目
田间管理按照常规,适时清理田间杂草。于3月21日追施肥水,施氮量为160 kg/hm2。每份品种材料于越冬期12月28日、1月6日,返青期2月25日去株行行头,分别随机取样10株调查苗期茎蘖数、叶绿素含量(即SPAD值,使用手持式叶绿素含量测定仪测定)并测量株高,于12月30日鉴定苗期春冬性习性,记录每份品种材料的拔节期、抽穗期、开花期及成熟期,小麦成熟期后调查记录定苗段内成穗数、穗粒数并于收获后测定千粒质量等产量构成因素。所有数据采用DPS生物统计软件进行统计和分析。
2结果与分析
2.1不同品种农艺性状的差异分析
将23个小麦品种的茎蘖数、叶绿素含量、苗期株高、拔节期、抽穗期、开花期、成熟期、千粒质量、成穗数及穗粒数等10个农艺性状项目分别进行品种间和区组间方差分析,表2结果表明,品种间的10个农艺性状差异均小于5%,农艺性状差异达显著或极显著水平,区组间仅茎蘖数、叶绿素含量、苗期株高差异极显著,其他性状均表现为不显著,因此综合分析可知这10个性状均适用于主成分分析。
2.2主成分分析
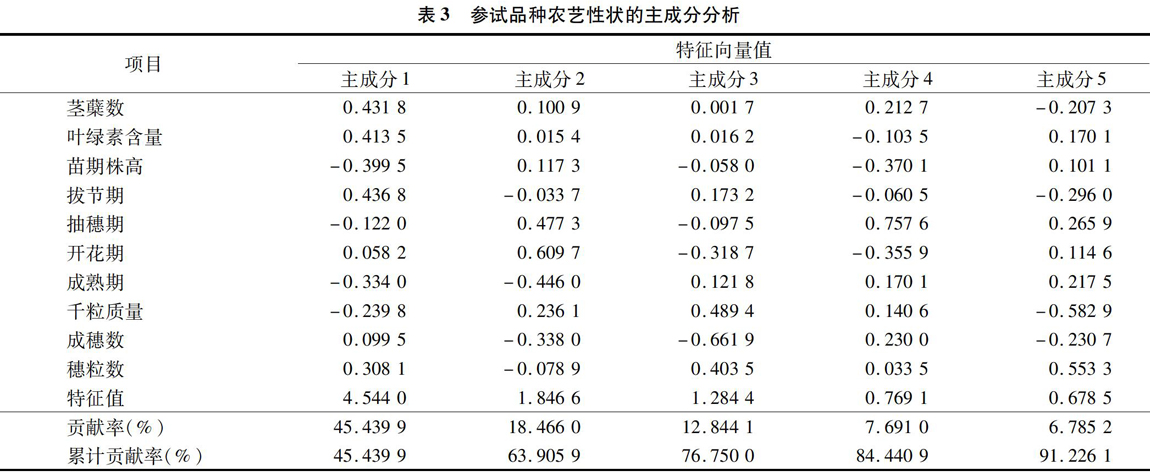
根据积累贡献率85%的标准选取5个主成分列出,表3结果表明,10个农艺性状中前5个特征值的累积贡献率达91.226%,说明10个农艺性状的绝大部分相关信息可由前5个主成分来概括。由表3还可知,第1主成分的特征值是4.544 0,贡献率高达45.44%。在第1主成分中以拔节期的特征向量值最大,其次是茎蘖数,说明农艺性状中拔节期对第1主成分的影响最大,其次是茎蘖数,因此将第1主成分称为拔节期生长因子,延长拔节期,能有效增加茎蘖数,提高叶绿素值,但苗期株高、成熟期、千粒质量依次降低或减少,说明在春冬性分类中,随着拔节期的延长,能明显增加茎蘖数,提高叶绿素值等冬性特性性状,但对苗期株高、成熟期、千粒质量起到一定的抑制作用。第2主成分的特征值为1.846 6,贡献率为18.466%。第2主成分中开花期的特征向量值最大,其次是抽穗期,所以将第2主成分称为开花期生长因子,由第2主成分的分向量值可以看出,开花期越长,抽穗期相应延长,千粒质量增加,成熟期缩短,成穗数减少,穗粒数亦有所减少,因此从产量构成角度来考虑,第2主成分均衡适中即可。第3主成分的特征值为1.284 4,所占权重为12.844%。第3主成分中成穗数的特征向量值最大(负值),说明成穗数对第3主成的影响最大,其次是千粒质量、穗粒数以及开花期(负值),因此将第3主成分称为成穗数产量因子,成穗数增加,开花期延长,千粒质量、穗粒数就相应减少,第3主成分值需在适量范围内为宜。第4主成分的特征值是0.769 1,贡献率为7.691%。第4主成分中抽穗期特征向量值最大,其次是苗期株高、开花期(均为负值),故将第4主成分称为抽穗期生长因子,第4主成分增大时,开花期提前,苗期株高降低。第5主成分的特征值是0.678 5,贡献率为6.785%。第5主成分中千粒质量的特征向量值最大(负值),其次是穗粒数,之后为拔节期(负值)、抽穗期、成穗数(负值)、成熟期;因此将第5主成分称为千粒质量产量因子;增大千粒质量,穗粒数减少,抽穗期缩短,拔节期、茎蘖数、成穗数则均有所延长或增加。
2.3参试品种的聚类分析结果
在主成分分析的基础上,以拔节期生长因子、抽穗期生长因子、开花期生长因子、千粒质量产量因子、成穗数产量因子这5个主成分因子为综合指标,依据欧氏距离,采用可变类平均距离法进行聚类分析(图1),在遗传距离为26.64的水平上把23份品种分为4类。以本试验鉴定结果为对照,其中第1类群包括龙麦33、石4185、衡观35、郑9023、扬麦16、宁麦13、扬麦25、西农979、小偃22、川麦104、绵麦367、云麦53,共12个品种;第2类群包括郑7698、周麦22、济麦22、烟农19、晋麦47、兰天26、新冬22,共7个品种;第3类群包括宁春4号、宁春50号、新春29,共3个品种;第4类仅包括强冬性品种肥麦。从聚类结果可以看出,第1类群除石4185、衡观35为半冬性品种,其余均为春性、弱春性品种;第2、第3、第4类群分别为冬性或半冬性、强春性、强冬性品种;其中宁春4号、宁春50号、新春29为强春性品种且都被分在第3类群;在4个类群中以第4类肥麦的冬性最强,生育期达357 d。由表4的原产地品种特性与本地(北纬33度线地区,下同)试验鉴定结果可知,23个品种中郑7698在原产地为弱春性,本地试验鉴定结果为半冬性;西农979在原产地为半冬性,本地试验鉴定结果为春性、弱春性,与本地试验鉴定对照表现出偏离,其余均与对照表现一致。这一特性表现也说明,各小麦生态区大面积主栽品种在本地具有较好的适应性和稳定性。
由表5对各类群农艺性状的平均值特征分析可以看出,第1类群中的拔节期、抽穗期、开花期均较早,成熟期适中,千粒质量降低,成穗数和穗粒数中等;第2类群中的拔节期较长,抽穗期、开花期最早,成熟期适中,千粒质量中等,成穗数较多,穗粒数适中;第3类群中的拔节期最早,抽穗期及成熟期最迟,开花期适中,千粒质量最高,成穗数和穗粒数最低;第4類群中的拔节期和抽穗期最迟,成熟期最早,千粒质量及成穗数适中,穗粒数最多。
3讨论
我国幅员辽阔,由于存在气候环境、栽培条件及耕作制度差异,各小麦生态区春冬性等小麦生育特性类型差异较大,本试验收集全国各生态区主栽品种23份,东北春麦区有黑龙江产地品种龙麦33,西北青藏春播麦区有产地品种肥麦,南方至西南麦区有川麦104、绵麦367、云麦53品种等。所选品种的地理远缘范围广泛,且为各地大面积主栽品种,品种遗传变异类型丰富,将原产地品种的春化特性与在本地鉴定的试验结果对比,发现仅西农979与郑7698表现有差异,其余对比结果均表现一致,印证了各小麦生态区大面积主栽品种在本地(北纬33度线地区)普遍具有较好的适应性和稳定性。
小麦的春化特性是小麦品种的重要性状之一,直接决定着小麦品种的种植范围和栽培措施,忽略小麦品种的春化特性进行引种、用种,往往会带来小麦贪青晚熟或造成冻害,给小麦生产造成巨大的经济损失[12]。目前研究者提出了许多冬春性小麦的鉴定方法和指标[13-14],鉴定方法主要有形态观察法、田间春播法[15]、人工模拟鉴定法(冰箱低温模拟春化)、综合顺序分类法[16]、分子生物学方法[15]等。其中,较普遍采用形态观察法,该方法简单、准确。本试验运用形态观察法进行小麦春冬性表型鉴定,并提取与春化特性有关的10个农艺性状进行主成分分析及聚类分析。聚类分析结果表明,本研究所用小麦品种可分为4个类群,分别是春性和弱春性类、冬性和半冬性类、强春性类、强冬性类,综合各农艺性状可知,春化生育特性相近的大多被分在1个类群,其中仅石4185及衡观35半冬性品种被分在春性类,究其原因,石4185及衡观35半冬性品种均是河北省大面积主栽品种,且品种春性特性相同,因此可能与其品种特性有一定关系,另根据莫惠栋等对大麦品种的聚类研究认为,用2~3个重要性状聚类,其分组结果清晰明确,便于材料取用,但其聚类稳定性差,而用多个性状聚类,其结果比较稳定,只是有些性状的差异会被另一些性状的差异掩盖,造成类群间差别模糊的结论[17]。因此聚类分析依据的性状、聚类方法等同样影响聚类结果[18],且与吴晓丽等的研究结果[19]相一致。综合分析可知,本研究分类结果较为准确、有效、可靠。
由主成分分析结果可以看出,第1主成分以拔节期特征向量值最大,作为小麦生育阶段重要标志的拔节期主成分权重达45.44%。金善宝等认为,小麦通过低温春化幼穗分化达二棱期标志春化阶段通过[20-21],谷冬艳认为,护颖分化期标志着春化效应的结束[22],说明由营养生长阶段转入快速生殖生长阶段,其表型形态与越冬苗期至返青起身期并最终进入快速生长拔节期的生育过程结果相一致[23]。由本研究结果看出,冬性越强,拔节越晚;春性较强,拔节较早。另据刘婷婷等的研究,单株茎蘖数越多,叶绿素含量越高,冬性越强[24],本研究表明,在10项农艺性状中分别以拔节期、抽穗期、开花期特征向量值最大的3个主成分值累计贡献率达71.6%,说明它们与春化特性关系密切,较客观地反映出在小麦快速生长发育阶段的表现,显著超越传统直观的单株茎蘖数、叶绿素含量、返青期株高等农艺性状因素的影响。另2项主成分中成穗数及千粒质量分别为12.84%和6.79%,占有一定比例,高于未入选穗粒数因素,这与刘婷婷等研究提出的在成穗数、千粒质量及穗粒数的产量构成三要素中,成穗数和千粒质量对产量的影响占主导作用的结论[24-25]相吻合。综上所述,10项农艺性状中5项主成分因子性状较能客观地反映其春化特性,特别以拔节期为主效应至抽穗期、开花期等快速生长阶段的表现更具反映春化发育的特性本质,这对于在小麦杂交育种亲本选配,尤其是聚合杂交最后1次回交所选配亲本材料的春化特性等重要农艺性状的评价有着重要的理论参考价值。
另外,种质资源春化特性分类研究可以与分子标记应用相结合,目前运用春化特性中感温性及感光性分子标记检测结果与表型鉴定结果不完全一致,但随着分子标记开发利用技术不断发展,与种质资源表型鉴定相结合进行育种改良仍是未来发展的主流趋势。
参考文献:
[1]Murphy J P,Cox T S,Rodgers D M,et al. Cluster analysis of red winter wheat cultivars based upon confficients of parentage[J]. Corp Science,1986,26(4):672-676.
[2]Brown J S. Principle component and cluster analysis of cotton cultivar variability across the US Cotton Belt[J]. Crop Science,1991,31(4):915-922.
[3]張学智,魏芝,扬珍,等. 春性硬粒小麦品种农艺性状的聚类分析[J]. 作物品种资源,1998,18(4):15-16,19.
[4]Butt M S,Anjum F M,van Z J,et al. Development of predictive models of end-use quality of spring wheats through canonical analysis[J]. International Journal of Food Science and Technology,2001,36(4):433-440.
[5]魏亦农,曹连莆. 二棱啤酒大麦品种资源农艺性状的聚类分析和主成分分析[J]. 种子,2003(3):69-70.
[6]胡立成,姚远,李秀兰,等. 黑龙江省大豆品种聚类分析初探[J]. 大豆科学,1991,10(1):10-16.
[7]Murty B R,Qadri M I. Analysis of divergence in some self-compatible forms of Brassica campestris var. Brown Sarson[J]. Genet,1966,26(1):4-58.
[8]蔡健,兰伟,罗瑞丽,等. 皖北小麦主栽品种遗传多样性的系统聚类法分析[J]. 中国农学通报,2006,22(11):143-146.
[9]张桂英,张国权,罗勤贵,等. 陕西关中小麦品质性状的因子及聚类分析[J]. 麦类作物学报,2010,30(3):548-554.
[10]刘二明,彭绍袭,黄费元,等. 水稻品种对稻瘟病抗性聚类分析[J]. 中国农业科学,1994,27(3):44-49.
[11]王林海,王晓伟,詹克慧,等. 黄淮麦区部分小麦种质资源农艺性状的聚类分析[J]. 中国农学通报,2008,4(4):186-191.
[12]苗果园,张云亭,侯跃生,等. 温光互作对不同生态型小麦品种发[CM(26]育效应的研究Ⅰ.品种最长、最短苗穗期及温光敏感性分析[LM]
[J]. 作物学报,1993,19(6):489-496.
[13]赵虹,胡卫国,詹克慧,等. 黄淮南片冬麦区主导品种春化基因及冬春性分析[J]. 西北植物学报,2010,30(3):495-504.
[14]梁宜策,米跃军,吕雪娥,等. 关于以小麦主茎全生育期的总叶片数量化品种冬春性的探讨[J]. 陕西农业科学,2011(1):53-57.
[15]王法宏,赵君实,荆淑民,等. 小麦不同类型品种在不同生态区的生育进程表现[J]. 莱阳农学院学报,1996,13(4):268-270.
[16]王建林,栾运芳,胡单,等. 栽培大麦冬春性判定的综合顺序分类法[J]. 大麦科学,2005(4):18-21.
[17]莫惠栋,顾世梁. 江浙沪大麦品种农艺性状的聚类分析[J]. 中国农业科学,1987,20(3):28-38.
[18]周丽艳,郭振清,马玉玲,等. 春小麦品种农艺性状的主成分分析与聚类分析[J]. 麦类作物学报,2011,31(6):1057-1062.
[19]吴晓丽,包维楷. 42份冬小麦品种主要农艺性状的聚类分析[J]. 西南农业学报,2010,23(4):1017-1022.
[20]金善宝. 中国小麦学[M]. 北京:中国农业出版社,1996:35-37.
[21]尹钧. 小麦温光发育研究进展Ⅰ.春化和光周期发育规律[J]. 麦类作物学报,2016,36(6):681-688.
[22]谷冬艳. 黄淮麦区小麦春化特性与播期效应研究[D]. 新疆:石子河大学,2007:33.
[23]赵广才,高德荣,常旭虹,等. 江苏省淮南地区小麦绿色高产高效技术模式图[J]. 麦类作物学报,2016,36(6):752-758.
[24]刘婷婷,王宝青,杨珍平,等. 九个黄淮和长江中下游冬麦区的优质冬小麦品种在晋中麦区的生育及品质表现[J]. 麦类作物学报,2015,35(2):182-191.
[25]刘孝成,石书兵,赵广才,等. 早熟型冬小麦群体性状及产量对氮磷肥和种植密度的响应[J]. 麦类作物学报,2016,36(6):752-758.
- 浅议“一支笔”财务审批制度
- 浅析电子企业现行成本法与多层次作业成本法核算的差异
- 会计信息失真的原因及其对策研究
- 基于我国财务会计规范改革中应当采用的最佳举措的思考
- 论联营项目的成本控制问题及对策
- 浅谈对企业财务影响的借款费用准则变化
- 作业成本法在我国民办高校教育成本核算中的应用
- 浅析存货跌价准备的核算
- 非货币性资产交换应用公允价值例解
- 论新制度下的高价互换配件的管理及核算
- 新准则下的固定资产会计计量属性
- 适应市场经济发展 提供准确会计信息
- 论企业成本管理创新
- 事业单位财务管理存在的不足及改进策略
- 厨房电器产品的定价策略
- 银行并购对我国银行业发展的价值分析
- 从产权入手解决我国公立医院的改革中的矛盾
- 我国服务贸易国际竞争力分析与对策
- 住房公积金管理初探
- 关于非货币资产交换当中商业实质判断的几点看法
- 浅谈国有建筑施工企业经济效益的影响因素及改进措施
- 对行政事业性国有资产管理模式的思考
- 工商行政管理系统对电子商务的监管对策分析
- 浅谈加强铁路职工社会保险档案管理工作的几点建议
- 大型美容化妆品连锁企业有效管理探讨
- marvelous
- marvels
- marvel²
- marvel¹
- marxism
- marxisms
- marxist
- marxists
- marzipan
- marzipanned
- marzipanning
- marzipans
- mas
- ma's
- masc
- mascara
- mascaraed
- mascaraing
- mascaras
- mascot
- mascotry
- mascot's
- observation
- observational
- observationally
- 篡乱
- 篡事
- 篡代
- 篡伪
- 篡位
- 篡位取代
- 篡位夺权
- 篡位而绝其后
- 篡位自立
- 篡党夺权
- 篡取
- 篡取君位
- 篡叛
- 篡国
- 篡夺
- 篡夺党和国家领导权
- 篡夺君位
- 篡夺君位或政权
- 篡夺君位而绝其后
- 篡夺君权之臣
- 篡夺国家政权
- 篡夺政权
- 篡夺权柄
- 篡夺的事
- 篡夺盗取