鱼类免疫系统的研究进展
张媛媛 宋理平
与哺乳动物不同,鱼类属于变温、低等动物。鱼类免疫系统主要由免疫组织与器官、免疫细胞和免疫因子组成,通常是指机体执行免疫应答和免疫功能的组织系统,是机体自身识别“自我”与消除“非我”,进而排除异己的一个功能体系。其主要功能有免疫防御、免疫监视及免疫自稳三方面。鱼类的免疫系统知识仍然在不断的完善过程中,与哺乳类一致,鱼类免疫也可分为特异性和非特异性免疫两大部分。一般来说,非特异性免疫在多数温度下都可以被激活,而特异性免疫如淋巴细胞的分化和抗体的产生更依赖于较高的温度。所以,鱼类更依赖于非特异性免疫系统来抵御病害的侵袭。目前,有关鱼类免疫系统上的研究多集中在免疫组织和器官、免疫受体、免疫因子以及免疫细胞等方面,现将现有的研究进行综述,以供参考。
1免疫组织与器官
免疫组织是机体发生免疫应答反应的场所。与哺乳动物类似,鱼类的免疫器官也包括中枢免疫器官和外周免疫器官。然而与哺乳动物不同的是,淋巴结和骨髓不属于鱼类的免疫器官,其免疫器官主要是指肾、脾、胸腺、消化道淋巴组织以及黏膜淋巴组织(MALT)等。其中,胸腺、肾脏和脾脏是鱼类最主要的免疫器官。
1.1胸腺
鱼类最重要的免疫器官主要是肾脏、脾脏和胸腺,而胸腺则被认为是鱼类的中枢免疫器官,其可通过促进淋巴细胞的增殖和相关激素的分泌等来促进机体的免疫调节功能。在免疫组织的发生过程中,胸腺可最先获得成熟的淋巴细胞,而后与头肾逐渐靠拢。胸腺主要分为内区、中区和外区,其中内区和中区分别类似于高等脊椎动物胸腺中的髓质和皮质,是鱼类T淋巴细胞成熟和分化的主要场所,并向外周血液释放成熟的T细胞。然而,荷尔蒙周期,外界环境的刺激,年龄的增长等或可导致鱼类胸腺发生退化,不同的鱼类其胸腺的寿命差异也很大,一些鱼类进入成鱼阶段,其胸腺就逐渐消失。卢全章的研究也证明了这一观点,草鱼在增长初期,胸腺发育较快,而到II龄以上其胸腺出现退化现象。
1.2头肾
肾脏是成鱼最重要的淋巴组织,其在长期的发育进化过程中可分为头肾(前肾)、中肾和后肾。头肾在鱼类的免疫应答过程中起主要作用,它可以自动分化成红细胞、淋巴细胞及各种粒细胞,是继胸腺后的第二个免疫器官。然而,并非所有鱼类均可产生粒细胞和B、T淋巴细胞。鱼的种类不同其所产生的免疫细胞也不同,有些鱼类仅产生B淋巴细胞,而有些鱼类可同时产生B、T淋巴细胞。魚类头肾残存于围心腔隔膜的前背侧,是一个沿着体轴呈Y型分离的器官,前端几乎垂直于体轴,是肾脏系统的一部分。其含有致密的血管网,包括许多造血组织和淋巴细胞丛,以及黑色素巨噬细胞(Melano-macrophages)和淋巴细胞。B淋巴细胞主要分布于鱼类头肾造血细胞和粒细胞生成的细胞群中,与黑色素巨噬细胞中心(MMC)和血管紧密相连,在免疫防御中起协同作用。头肾无肾小球,失去排泄功能,主要负责造血、异源物质吞噬、IgM的形成和免疫记忆功能。另外,头肾也是一个内分泌器官,类似于哺乳动物的肾上腺,会释放皮质醇和一些其他的激素。在一定程度上,我们可根据头肾的重量、大小以及发育状况来判断鱼类的生长发育状况,前人的许多研究表明,鱼类头肾的重量与鱼体的年龄呈正相关,与鱼体的增重率也紧密相连。此外,与哺乳动物淋巴结相类似,鱼类肾脏可直接有效地抵御外界病原的入侵,不依赖因抗原刺激而产生相应的免疫细胞便可有效抵御外界病原的入侵。
1.3脾脏
一般说来,脾脏分为红髓和白髓,但只存在于软骨鱼中,硬骨鱼却没有,这说明只有软骨鱼才具有真正意义上的脾脏结构,但不论脾脏有无真正的白髓和红髓之分,其都具有免疫功能。脾脏是红细胞和各种粒细胞等产生、储存和成熟的主要场所,可为鱼类机体提供充足的血液和大量免疫细胞。在硬骨鱼类中通常只有一个脾脏,其位于胃或肠附近,是一个形态结构比较固定的淋巴组织。其功能主要包括:大分子物质的清除,抗原的降解和加工以及抗体生成。脾脏也是主要的造血器官,能产生血细胞、内皮细胞、网状细胞、巨噬细胞和黑色素巨噬细胞。随着鱼体的生长与发育,其免疫器官也随之发生巨大变化。成年软骨鱼的脾、肾较之硬骨鱼有很大差异,其肾脏失去了造血功能,脾脏则成长为机体的重要淋巴器官,而硬骨鱼则相反。张永安等研究表明,当硬骨鱼受到一定的外源刺激时,其脾、肾等免疫器官可产生大量的黑色素巨噬细胞,进而可通过与抗体和淋巴细胞相结合来调节机体免疫机能。
1.4黏膜相关淋巴组织
黏膜相关淋巴组织(MALT)是一种主要分布于机体黏液组织中的淋巴细胞生发中心,因其不具备完整的淋巴结构,故而被称为黏膜相关淋巴组织。它是鱼类除以上免疫器官外的又一重要免疫器官。巨噬细胞和淋巴细胞等广泛存在于鱼类的鳃、皮肤和消化道等的上皮组织中。当机体受到外源因子刺激时,淋巴细胞会产生大量的抗体分子与被巨噬细胞处理的抗原发生特异性结合,并与机体中其他免疫因子一起促进机体免疫能力的提高,保护机体不受病原菌侵害。
2鱼类细胞免疫
能够参与免疫应答或与免疫应答相关的细胞,均可被称为免疫细胞。主要分布于机体淋巴液、血液及各种免疫器官和组织中。免疫细胞的分布及含量在一定程度上反映了机体免疫功能的强弱,其主要包括可吞噬大量外源微生物,在机体中参与非特异性免疫反应的吞噬细胞和参与机体特异性免疫反应的淋巴细胞。二者在机体免疫系统的调节中均起重要作用。
2.1淋巴细胞
淋巴细胞(Lymphocyte)是机体免疫应答的重要细胞成分,主要由淋巴器官产生,是白细胞的一种。在哺乳动物中,淋巴细胞可分为T、B淋巴细胞两个亚群,二者表面具有不同的抗原和受体,在机体中参与不同的免疫反应。其中,B细胞主要分泌抗体,参与机体特异性免疫反应;而T细胞可通过分泌细胞因子来调节机体免疫功能,也可直接杀伤细胞,在机体中主要参与非特异性免疫反应。在硬骨鱼类淋巴细胞中,能表达表面膜免疫球蛋白(surface membrane immunoglobulin,smIg)的为B淋巴细胞,不能产生免疫球蛋白(Ig)的为T细胞。近年来,单克隆抗体、流式细胞仪和分子生物学等新型技术的广泛应用使我们对鱼类T、B细胞的认识不断地深入。一般来说,我们可采用单克隆抗体技术来区分淋巴细胞表面抗体受体的活性,然而到目前为止,单克隆抗体技术还不能区分T细胞。Miller等对鲶鱼免疫细胞进行了研究,结果显示单抗可与鱼体T细胞发生反应,但同时又与凝血细胞和中性粒细胞发生交叉反应,因此鲶鱼中是否存在T淋巴细胞尚需要更进一步的研究证实。但鱼类至少具有功能性的淋巴细胞是毋庸置疑的。同时也有研究证明,鲤鱼胸腺内的T淋巴细胞含量最多且不随孵化时间的增长而变化,而其他免疫器官中的T淋巴细胞逐渐减少甚至消失;然而B淋巴细胞在孵化后两周最先出现在鲤鱼头肾中,机体的其他器官中除脾脏和血液外B细胞含量均较少。此外,大菱鲆(Scophthalmus maximus)和海鲈(Lateolabrax.japonicus)的肠黏膜和黏膜下层中均发现较多的T细胞,而B细胞主要在固有层。
2.2吞噬细胞
鱼类吞噬细胞是组成非特异性防御系统的重要成分,主要包括单核细胞、巨噬细胞和各种粒细胞。其在机体的防御过程中均起到不可替代的作用。当机体受病原菌感染时,各组织中的吞噬细胞会对外源因子进行识别及吞噬并有效降解和摄取微生物及其产物,阻止其进入机体循环系统,从而对机体进行保护防止病原菌感染。同时,吞噬细胞还可分泌大量细胞因子对机体进行免疫调控。
单核细胞在动物体的非特异性免疫中起关键作用。与哺乳动物相似,鱼类的单核细胞具有很强的黏附及吞噬能力,可大量吞噬进入机体的外源微生物及本身受损、凋亡的细胞。单核细胞是一类普遍存在于所有硬骨鱼血液中并由造血细胞分化而成的终末细胞。其可在适宜环境中随血液进入各组织器官并发育成为不同的巨噬细胞群。另外,徐豪等研究证明,环境污染以及寄生虫和细菌等外源物质感染均可使鱼体血液中单核细胞增多,这一现象可能与鱼类内分泌系统有关。
在硬骨鱼类中,脾脏和肾脏是生成粒细胞的主要场所。然而根据颗粒染色不同粒细胞有嗜中性、嗜碱性及嗜酸性之分。嗜中性粒细胞是先天性免疫中的一种主要效应细胞,其可通过胞吐作用对外来微生物进行识别并破坏。当机体产生炎症反应时,其细胞内含有的大量杀菌剂及潜在毒性物质被释放到胞外并配合发生脱粒反应。中性粒细胞在炎症发生时的功能仍存在很大争议,一些报道认为其可产生大量免疫调控酶对机体进行免疫调控。另外,当鱼类遭受外界刺激后,其嗜中性粒细胞也显示出化学发光性和趋化性。
鱼体中并非都含有嗜酸性和嗜碱性粒细胞。大多数鱼类仅含有嗜酸性粒细胞,少部分鱼类才含有嗜碱性粒细胞,然而还有几少部分鱼类中二者均不存在。有研究证明,在大马哈鱼(On-corhynchus keta)中可以检出嗜酸性粒细胞和嗜碱性粒细胞,然而在鲫鱼(Carassius auratus)中,却没有观察到嗜碱性粒细胞。这可能是由于嗜碱性粒细胞含量较少且在制片过程中极易被分解,导致其很难被观察到。嗜酸性粒细胞由造血淋巴器官产生,其可随血液循环进入不同器官中,随之分化为粒细胞。与哺乳动物肥大细胞相似,鱼类机体在遭受病菌感染和外界刺激時,其嗜酸性粒细胞可发生脱粒反应并释放细胞内活性物质及合成、分泌大量免疫分子和相关免疫调控酶,从而调节机体免疫功能。此外,嗜酸性粒细胞也具有吞噬功能,当机体遭遇寄生虫长期的感染时,其可聚集在寄生部位发挥吞噬作用以抵御寄生虫的侵袭。
鱼类的巨噬细胞种类繁多,当机体遭受细菌和寄生虫感染时,可分泌多种细胞因子和二十碳四烯酸等以抵御病原菌感染。当鱼体遭受病原菌感染时,巨噬细胞发生吞噬作用,会引起呼吸爆发,同时产生肿瘤坏死因子(tumor necrosis fac-tor,TNF)。此外,机体受到外源刺激时,巨噬细胞可通过识别并传递抗原,调节淋巴细胞活性及控制自身的增殖与分化等途径来对机体免疫系统进行调控。许多研究证明,当鱼体遭受外源刺激时,其吞噬细胞可大量吞噬异物并分泌活性物质与巨噬细胞、MMC共同形成聚集体,以保护机体不受损伤。
2.3细胞吞噬作用及活性氧代谢
吞噬细胞表面受体可与病原菌(或其自身产生的趋化因子相结合,激活其酯酶和HMP途径,使细胞内Ca2+大量流失,导致微丝和微管装置推动细胞向病原菌的方向迅速移动,称为趋化作用。而调理作用是指抗体和补体等调理素与病原体相结合或覆盖于病原体的表面,通过与巨噬细胞表面Fc受体或补体受体结合,从而促成吞噬细胞对病原菌的吞噬作用。当吞噬作用被激活,将会产生一系列的代谢产物,如活性氧(Reactive oxygenspecies,ROS)、一氧化氮(Nitric oxide,NO)、相关酶和细胞因子(Cytokine)。
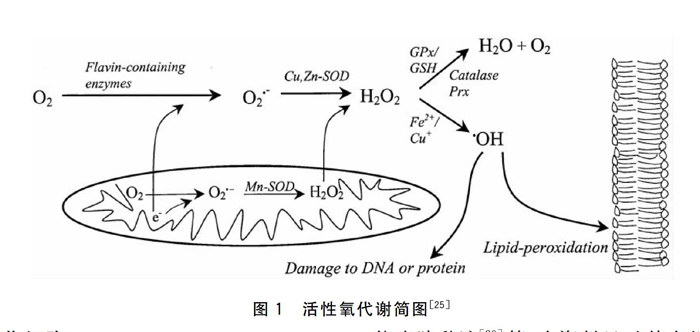
在巨噬细胞、单核细胞和粒细胞等吞噬细胞的吞噬过程中,将会产生大量的活性氧,这是杀灭微生物的一个重要手段12引。吞噬细胞在此过程中会消耗大量氧气,产生大量的超氧化物阴离子(superoxide anion,O2-),所以称之为呼吸爆发(Respiratory burst)。O2-的产生受制于一种酶,它与哺乳动物吞噬细胞中的NADPH氧化酶相似,可与细胞膜上的受体相结合。O2-氧化活性不高,不能透过膜。然两分子的O2-被超氧化物歧化酶(superoxide dismutase,SOD)歧化成过氧化氢(Hydrogen peroxide,H2O2)。过氧化氢则可以通过膜,其代谢途径有:由嗜中性粒细胞中溶酶体分泌的髓过氧化物酶(myeloperoxidase,MPO)催化产生次氯酸(Hypochlorous acid,HOCl);在过氧化氢酶(Catalase,CAT)或谷胱甘肽过氧化物酶(Glutathione peroxidases,GPx)催化下形成水;通过金属离子(Fe2+或Cu+)形成羟基阴离子(Hydroxyl radical,·OH)。其中,HOCl和·OH是两种对细菌有高毒性的活性氧分子,可在机体中用作杀菌。·OH也是引起脂质过氧化(1ipid peroxidation)和DNA或蛋白损伤(Damage to DNA or protein)的重要分子。详见图1。
2.4自然杀伤细胞
鱼类中的自然杀伤细胞被称为非特异性毒性细胞(Non-specific cytotoxic,NCC),是最小的白细胞,呈多形性,细胞核有裂口,细胞质相对较少,细胞表面有微绒毛,其可通过长的胞膜纤维吸附于靶细胞上;且鱼类的头部、脾肾及末梢血管中的NK细胞可直接杀伤鱼体中各种靶细胞。NK细胞的杀菌作用是自发的,不需要任何的诱导,这类细胞在机体中与靶细胞接触后,可通过自身产生淋巴毒素来杀伤和破坏靶细胞。在软骨鱼中,这种具有自发杀菌作用的细胞被普遍认为是巨噬细胞的一种。而硬骨鱼中的这类细胞与哺乳动物NK细胞不同,它是一种较小的淋巴细胞样细胞。其胞液中不含颗粒物质,呈多种晶型,可通过参与细胞毒性作用来调节机体免疫功能。
3鱼类免疫分子
鱼类机体中同时存在特异性和非特异性免疫系统,特异性免疫是指当机体受抗原刺激时,淋巴细胞发生分化与增殖,并产生免疫球蛋白的应答过程。而非特异性免疫反应是机体与生俱来的一种固有免疫反应,其两套系统都由不同的体液免疫分子参与介导免疫反应。这些免疫因子构成了机体正常免疫功能的分子基础。
3.1抗菌肽
鱼类体表及外层可分泌大量分泌物,如多功能皮肤黏液等,有资料显示其在机体防御细菌及病毒等免疫过程中起重要作用。在动植物及哺乳类动物中,抗菌肽被作为先天性免疫反应中的一个因子而广泛存在。其氨基酸数小于100,均为碱性多肽,常带正电荷,N端有多聚腺苷酸残基,C端均被酰胺化,第二位氨基酸多为色氨酸,且具两亲性二级结构。其抗菌机理主要是在细菌细胞膜上形成离子通道,破坏膜电位,从而引起细胞内容物泄漏,进而杀灭细菌。其对革兰氏阴性菌和革兰氏阳性菌均有抑菌效果。当鱼体受到损伤或微生物刺激时,机体能够迅速产生抗菌肽以抵御病原菌及微生物的侵袭。
3.2补体
鱼类补体系统与高等脊椎动物一样可分为三个补体激活途径:经典途径(CCP),替代途径(ACP)和凝集素途径(LCP)。补体途径是由一组30种以上的可溶性糖蛋白(包括补体分子C1-C9以及一组主要与旁路途经有关的分子,如B因子和D因子等)组成。经典途径是指被细胞表面的抗体激活,包括一些急性蛋白激活及病毒、细菌或病毒感染细胞激活;替代途径不依赖于抗体,其直接可由病毒、细菌、真菌或肿瘤细胞等外来微生物激活;凝集素途径的激活是由结合在微生物细胞表面的蛋白复合体(甘露糖结合凝集素,mannose-binding lectin,MBL)、与MBL相关的蛋白酶和与MBL相关的小蛋白,以碳水化合物群的形式结合在细菌的表面,丝氨酸蛋白酶(MASPs)和葡萄糖结合凝集素(GBL)替代经典途径中抗原抗体复合物,在不依赖抗体的情况下,激活凝集素途径。然而,有关硬骨鱼类中MBL关联丝氨酸蛋白酶的编码基因的分子研究和作用机制却鲜为人知。所有这三种途径收敛到裂解途径,从而引起调理素作用或者直接导致微生物死亡。
3.3抗体
抗体是指当机体在遭遇抗原刺激下,由记忆细胞或B淋巴细胞增殖分化而成的浆细胞所产生的一类具有免疫功能的、可与相应抗原发生特异性结合的球蛋白。其主要分布在血清中,也分布于组织液及外分泌液中。世界卫生组织在1964年将免疫球蛋白(Ig)统一定义为具有抗体活性以及与抗体相关的球蛋白。一般来说,免疫球蛋白包括已证明抗体活性的正常免疫球蛋白和未证明抗体活性的异常免疫球蛋白(如异常Y球蛋白血症患者血液中的免疫球蛋白)。因此,所有抗体均为免疫球蛋白,而并非所有的免疫球蛋白均为抗体。
整个抗体分子可分为两部分,即恒定区和可变区。在同一物种中,不同抗体分子的恒定区都具有相同或几乎相同的氨基酸序列。哺乳动物可根据重链不同将抗体分为五种不同类型的免疫球蛋白(IgM,IgA,IgD,IgE,IgG)。在硬骨鱼中,迄今为止,仅发现IgM和IgD两类免疫球蛋白。免疫球蛋白的主要功能包括与特异性抗原(包括外来的和自身的)相结合从而有效地清除侵入机体内的微生物、寄生虫等异物,激活补体与Fc受体相结合从而激发调理作用。鱼类与哺乳动物相比,鱼类抗体形成的时间长,抗体滴度低。抗原(包括抗原性质、剂量、免疫方式等)、机体状态(性别、年龄、体重、营养状况及激素水平等)和环境变化(胁迫、温度、光照及水体中有机物和无机物等)都会影响鱼类免疫球蛋白表达水平。
3.4细胞因子
在先天性非特异性免疫中,机体释放了大量的促免疫细胞因子。在过去的几年里,有关于鱼类趋化因子和细胞因子的研究越来越多。细胞因子是一类由外源微生物及其他刺激剂诱导多种免疫细胞产生的可溶性蛋白质,其分子量低,具有调节机体特异性及非特异性免疫、促进血细胞分化与增值、修复受损细胞等功能。细胞因子主要分为白介素、干扰素、肿瘤坏死因子、生长因子及趋化因子等,其可通过与靶细胞表面受体相结合并将刺激信号传人机体细胞,从而调节细胞的增殖与分化,进而对机体免疫功能进行调控。同时,免疫因子还可通过各种分泌方式如自分泌等调节机体免疫机能和生理代谢反应。
3.4.1肿瘤坏死因子 在哺乳动物中,肿瘤坏死因子主要由活化的巨噬细胞、单核细胞、中性粒细胞、自然杀伤细胞和T细胞分泌产生,能杀伤和抑制肿瘤细胞,促进中性粒细胞的吞噬作用,诱导肝细胞急性期蛋白的合成以及促進细胞的增殖和分化,是一个重要的炎症因。TNF-α主要由单核巨噬细胞分泌,TNF-β主要由T淋巴细胞分泌。大量研究证明TNF-α是一个由白细胞分泌的重要的巨噬细胞激活因子(macrophage-activating factor,MAF)。且有研究显示鱼体中多发现TNF-αmRNA的表达。在虹鳟(Oncorhynchus mykiss)、大菱鲆(Scophthalmusmaximus)、海鲤(Nemadactylus bergi)中,均发现MAF能激发巨噬细胞的活性反应,提高呼吸爆发活性、吞噬作用和一氧化氮的含量。
3.4.2干扰素(Interferons,IFNs) 干扰素是一种分泌蛋白,能够在转录水平上调节干扰素刺激基因(IFN-stimulated genes,ISGs)的表达,进而使脊椎动物的细胞进入一种抗病毒的状态。在哺乳动物中,根据基因序列、蛋白结构和功能的不同将干扰素分为三种不同的类型,分别是type I IFN、type II IFN和IFN-λ。 Type IIFN包括IFNα和IFNβs,在大部分细胞中主要由病毒诱导分泌。然而Type II IFN与IFN-λ均是由自然杀伤细胞和T淋巴细胞分泌产生。在斑马鱼(Batchydanio rerio war.)、大西洋鲑鱼(Salmo salar)、金娃娃鱼(Tetraodon nigroviri-dis)等中也克隆出了干扰素基因。干扰素的作用机制为当细胞感染后分泌的干扰素能够与周围未感染细胞上的相关受体作用,促使这些细胞合成抗病毒蛋白防止进一步的感染和复制,从而起到抗病毒作用。在哺乳动物中,抗病毒作用是由干扰素刺激基因诱导产生的蛋白所引起的,这是一种黏液病毒蛋白(myxovirus,Mx),存在于从哺乳类到鱼类的细胞质或细胞核中,具有抗病毒的活性。它也是硬骨鱼类中唯一报道的能由干扰素产生的抗病毒蛋白。
3.4.3白介素 白细胞介素(Interleukins,IL)简称白介素,是一类由白细胞等多种细胞产生并具有重要调节作用的糖蛋白。它属细胞因子,在机体中可与血细胞生长因子互相协作、相互协调,共同完成免疫调节和造血功能。IL在信息的传递,免疫细胞的激活与调节,T、B淋巴细胞的增殖与分化以及炎症反应中均起重要作用。IL-1主要由巨噬细胞产生,此外几乎所有的核细胞,如B细胞、NK细胞、体外培养的T细胞、中性粒细胞及平滑肌细胞均可产生IL-1。它是促炎症因子的一种,能够通过调节其他细胞因子从而间接引起一连串的反应,进而导致炎症。IL-1主要分为IL-1α和IL-1β两种不同的分子形式,但他们结合相同的细胞表面受体,发挥相同的作用。在硬骨鱼类的虹鳟和大西洋鲑中也发现了IL一1β的受体,其在鱼体头肾、脾脏、鳃中表达较高,在炎症反应中起重要作用。
3.4.4趋化因子(Chemokines) 趋化因子是一类低分子(多为8—10KD)的细胞因子家族蛋白(如IL-8、MCP-1等),其具有定向细胞趋化作用。其在机体中既可促进机体炎症反应,又可控制机体在正常的修复和发育过程中的发生的细胞迁徙,对机体的免疫调控具有重要作用。有一些趋化因子可控制机体免疫细胞趋化反应,如诱导淋巴细胞到淋巴结等。这些淋巴结中的趋化因子可通过与这些组织中的抗原提呈细胞相互作用而监视病原体的入侵。趋化因子存在于所有的脊椎动物与一些病毒及细菌中,而在无脊椎动物中则不存在。许多研究证明,在硬骨鱼类的虹鳟(On-corhynchus mykiss)、鲤鱼(Cyprinuscarpio)、鲶鱼(Silurus asotus)及比目鱼(Paralichthys olivaceus)等中均确定有趋化因子的存在。
3.4.5金属离子结合蛋白 金属离子结合蛋白包括转铁蛋白、血浆铜蓝蛋白和金属硫蛋白。血清转铁蛋白(Transferrins,Tf),又称为铁传递蛋白,是鱼体内铁的运输者,是血清中一种非血红素结合铁的β-球蛋白。不同种属的鱼类虽具有不同的Tf,但每个Tf均含有2个三价铁离子的结合位点。铁是微生物代谢和繁殖所必需的基本元素,由于转铁蛋白在机体中具有很高的铁结合能力,其可将机体吸收及储存的铁离子运输到红细胞供给血红蛋白使用,从而防止机体铁元素的损耗。同时,转铁蛋白对微生物的分化与增值均具有抑制作用;血浆铜蓝蛋白(Ceruloplasmin)又称为亚铁氧化酶,是一种由单一多肽链组成的蓝蛋白,其可以螯合铜和其它的二价阳离子。例如它可以将亚鐵离子氧化为铁离子,然后将铁离子结合到转铁蛋白上,促进机体对铁元素的吸收及利用。同时,血浆铜蓝蛋白还可加速铁元素的运输,使其得到充分利用;金属硫蛋白(Metallothio-nein,Mt)分子量低且热稳定,其含有丰富的可以与铜、锌、钙和汞等金属离子相结合的半胱氨酸。当机体遭受外源因子刺激或发生炎症反应时,可产生大量金属硫蛋白与相应细胞膜上受体发生特异性结合,直接或间接影响机体吞噬能力及呼吸爆发活性,从而调节机体免疫功能,杀灭病原微生物。
4小结
总的来说,进入二十一世纪以来有关鱼类免疫学研究取得了很大进步。同时,也引起了众多免疫学家的广泛重视,对鱼类免疫系统的进化及鱼病的防治提供了一定的理论指导。然而,鱼类免疫系统错综复杂,目前对鱼类特异性免疫和非特异性免疫机制的研究并不完善。尤其是如何选择一种天然、无毒副作用的药物防治鱼病、提高机体非特异性免疫力,同时也不影响健康鱼体的生长,这还需要进一步的研究与探索。
(收稿日期:2017-12-05)
