酵母多糖、壳聚糖、甘草酸对草鱼非特异性免疫功能的影响
张涛


摘 要:将草鱼放入网箱中,在基础饲料中分别添加不同剂量的酵母多糖、壳聚糖和甘草酸后,进行投喂,研究其对草鱼血清中菌酶(LSZ)、过氧化氢酶(CAT)及补体C3活性的影响。结果表明:投喂添加中药免疫添加剂的饲料49 d 后,与对照组比较,酵母多糖添加剂各组、壳聚糖高剂量组草鱼血清LSZ活性显著提高(P<0.05);酵母多糖低剂量组、中剂量组和壳聚糖高剂量组草鱼血清CAT含量显著提高(P<0.05);酵母多糖中剂量组草鱼血清补体C3活性显著提高(P<0.05),说明中药免疫添加剂对增强草鱼免疫活性具有一定的作用。
关键词:中药免疫添加剂;草鱼;LSZ;CAT;补体C3
草鱼俗称鲩鱼、鲩,东北俗称草根子,属鲤形目,鲤科,雅罗鱼亚科,草鱼属,为中国东部广西至黑龙江等平原地区特有经济鱼类;其生长迅速,饲料来源广,肉质鲜美,是中国淡水养殖的四大家鱼之一。近年来,随着我国淡水养殖业的发展,人工养殖草鱼受到水环境、饲料及养殖管理等条件变化因素影响,常常发生疾病而造成大量死亡,严重损害了我国淡水养殖业的发展。因此,研制草鱼免疫添加剂,促进鱼体的疾病抵抗能力,对人工养殖草鱼技术的发展具有极其重要的意义。
免疫添加剂因为具有高效、毒副作用少、无污染等特点,是抗生素和化学药物的最佳替代品,因此受到水产工作者的亲睐。近年来,由国内外的研究报道可知,水产动物饲料中添加免疫添加剂可促进水产动物体的非特异性免疫指标的提高[1-5],从而增强水生动物的免疫能力;曹振杰等[6]在草鱼饲料中添加不同剂量的免疫多糖明显增强了鱼体的免疫能力,提高了SOD活力。本试验中,在草鱼的饲料中添加3种不同的免疫添加剂饲喂草鱼,并通过试验数据分析草鱼血清中的几种非特异性免疫指标,其目的是考察草鱼的饲料添加剂能否有效提高草鱼非特异性免疫指标,从而增强草鱼抵抗疾病的能力。
1 材料与方法
1.1 试验方法与饲料
在草鱼的基础饲料中分别添加相同浓度梯度的酵母多糖、壳聚糖和甘草酸,浓度梯度分别设计为0.22%、0.44%、0.66%,共9试验组,一个对照组,每组设3个重复。饲料配制时,饲料原料经粉碎过40目筛后,制得3 mm颗粒,草鱼基础饲料配方见表1。
1.2 试验用鱼及养殖管理
试验草鱼平均体重97.6±5.2 g,购自辽宁省淡水水产科学研究院试验场,暂养后挑取相同规格的健康草鱼750尾,随机分为10组,每组3个平行,即1个对照组和9个试验组,共计30个网箱,网箱大小为1.2 m×0.6 m×1.2 m,放在8.5 m×1.6 m×8.5 m的水泥池塘中,每个网箱随机放入25尾草鱼。
试验期间水温26±2 ℃,每天在7∶00、12∶00、17∶00投喂饲料,投饲量为鱼体重的4%,连续充氧,每日排污和换水各一次,共计49 d;其中,养殖水体NH+4-N<0.5 mg/L ,pH(7.4±0.3),DO≥6 mg/L。
1.3 血清免疫指标的测定
试验结束后,从每个试验箱中取6尾草鱼,尾静脉采血,3 000~5 000 r/min 离心 8 min,取血清于5 ml离心管中备用。溶菌酶(LSZ)、过氧化氢酶(CAT)、补体C3活性均采用由南京建成生物技术研究所生产的试剂盒的测定方法测定。
1.4 数据分析与处理
试验结果用“平均值±标准差”表示,采用SPSS14.0在a=0.05水平下进行统计分析,数据经单因素方差分析后,进行Duncans 多重比较统计分析。
2 结果
2.1 三种免疫添加剂对草鱼血清溶菌酶(LSZ) 活性的影响
由表2和图1可知,草鱼血清溶菌酶(LSZ)活性由高到低依次为酵母多糖中剂量组、酵母多糖低剂量组、壳聚糖高剂量组、酵母多糖高剂量组、壳聚糖中剂量组、壳聚糖低剂量组与甘草酸中剂量组、甘草酸高剂量组、甘草酸低剂量组;其中,酵母多糖添加剂各组、壳聚糖高剂量组与对照组比较,草鱼血清LSZ活性均显著提高(P<0.05);其他各组与对照组比较差异不显著(P>0.05),只有甘草酸低剂量组草鱼血清LSZ活性略低于对照组。
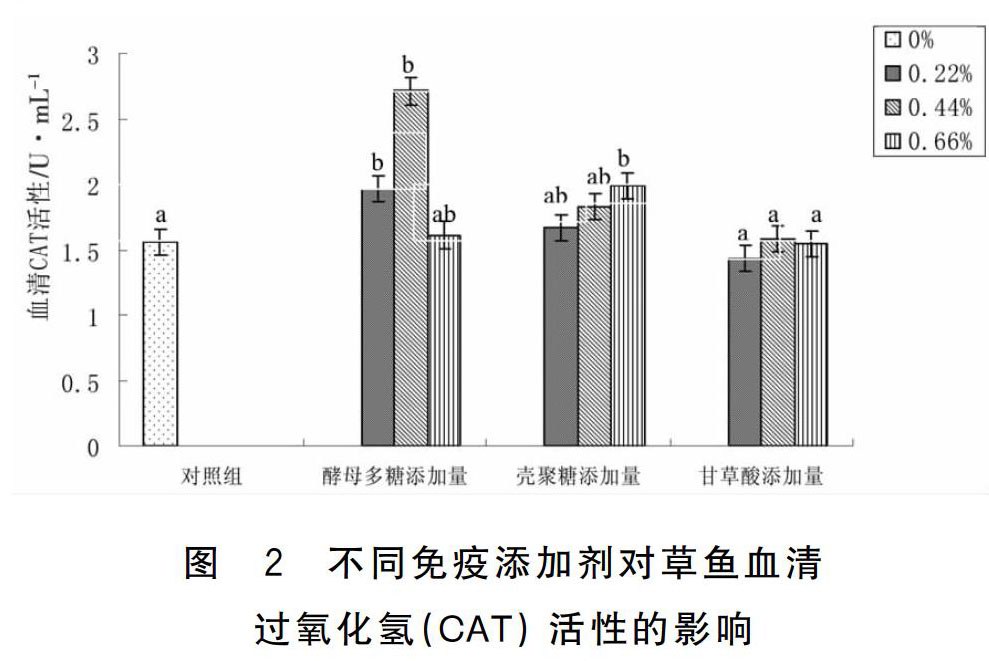
2.2 三种免疫添加剂对草鱼血清CAT活性的影响
由表2和图2可知,酵母多糖低剂量组、中剂量组和壳聚糖高剂量组草鱼血清CAT含量显著高于对照组(P<0.05),其余各组与对照组比较差异不显著(P>0.05),只有甘草酸低剂量组与高剂量组草鱼血清CAT略低于对照组。
2.3 三种免疫添加剂对草鱼血清补体C3活性的影响
由表2和图3可知,酵母多糖中剂量组草鱼血清补体C3活性显著高于对照组(P<0.05),其余各试验组与对照组比较呈不显著差异(P>0.05),只有甘草酸低剂量组草鱼血清补体C3活性略低于对照组。
3 讨论
鱼类作为低等脊椎动物,其非特异性免疫功能具有抵抗外来病原入侵的主要作用[7-9]。
溶菌酶(LSZ)是一种能水解致病菌中黏多糖的碱性酶,该酶具有抗菌、消炎、抗病毒等作用。溶菌酶(LSZ)作用机理主要是通过对菌体细胞壁进行破坏而使不溶性黏多糖分解成可溶性糖肽,导致菌体细胞壁破裂,胞壁里内容物质逸出而使菌体溶解[10]。另外,溶菌酶可与病毒蛋白结合,并与RNA、DNA、脱辅基蛋白形成复合盐,致使病毒失去活性。近些年研究发现,中草药的免疫添加剂对水产动物LSZ具有增强作用。胡宗云[11]等研究發现饲料中添加硬葡聚糖在增强草鱼血清溶菌酶(LSZ)活性方面效果更好;许国焕[12]等研究发现肽多糖能提高彭泽鲫血清溶菌酶活性;王树芹[13]等在异育银鲫饲料中添加0.5%或1.0%的壳聚糖能够显著提高溶菌酶活性(P<0.01),这与本研究添加0.66%壳聚糖的饲料能显著提高草鱼血清溶菌酶(LSZ)活性(P<0.05)相近;同时,本研究在饲料中添加了0.22%、0.44%、0.66%的酵母多糖后,草鱼血清溶菌酶(LSZ)活性均显著提高(P<0.05),这与李桂峰[14]等研究发现添加了0.5%和0.8%酵母多糖的饲料能显著提高赤眼鳟溶菌酶活性(P<0.05)相近。
过氧化氢酶(CAT)是一种酶类清除剂,可清除机体氧自由基,是生物防御体系的重要抗氧化物关键酶;当鱼类体内过氧化氢酶(CAT)活性降低时,鱼体内自由基过多会使其代谢混乱、内分泌失调、机体免疫能力下降等,从而使鱼体易受到病原微生物入侵,最终导致鱼体发病[15-16]。王洪斌[17]等研究发现日本蟳在注射海洋红酵母多糖48 h后,其血清中的过氧化氢酶(CAT)提高35%;这与本试验中饲料在分别添加了0.22%、0.44%的酵母多糖后,草鱼血清过氧化氢酶(CAT)活性显著提高(P<0.05)相近。目前,关于鱼类饲料中添加壳聚糖能够使鱼类血清CAT活性增高未见报道;但本试验中饲料在添加了0.66%的壳聚糖后,草鱼血清过氧化氢酶(CAT)活性显著提高(P<0.05)。补体C3是机体抵抗微生物入侵的重要组成部分,是一个具有精密调控机制的蛋白质反应系统,并由多种具有酶活性的球蛋白构成,其主要存在机体的血清、组织液中;当该系统通过一定途径被激活时,可发挥调理吞噬、裂解细胞、免疫调节等功能,这在鱼类早期抗感染过程中发挥着重要作用[11]。李桂峰[14]等研究发现添加了0.5%和0.8%酵母多糖的饲料能显著提高赤眼鳟补体C3水平(P<0.05);这与本试验中饲料在添加了0.44%的酵母多糖后,草鱼血清补体C3活性显著提高(P<0.05)相近。于翔[18]等研究发现添加一定量壳聚糖的饲料能显著提高草鱼血清补体C3水平(P<0.05),这与本试验饲料添加了0.22%、0.44%、0.66%的壳聚糖后,草鱼血清补体C3活性提高相近;但本试验提高补体C3水平不显著(P>0.05)。
刘丽萍[19]等报道甘草酸是有效的免疫添加剂,其免疫调节作用表现在对免疫活性细胞、细胞因子、补体等多方面。而本研究在饲料中添加了 0.22%、0.44%和0.66%的甘草酸后,草鱼血清溶菌酶(LSZ)、过氧化氢酶(CAT)、补体C3活性与对照组差异都不显著(P>0.05),低剂量组的溶菌酶(LSZ)、过氧化氢酶(CAT)、补体C3活性甚至低于对照组,笔者认为这是甘草酸并不能有效激活草鱼血清中的溶菌酶(LSZ)、过氧化氢酶(CAT)、补体C3活性所致。
参考文献:
[1]
Chen S C,Yoshida T,Adams A,et al.Non-specific immune response of Nile tilapia, Oreochromis nilotiea, to the extracellular products of Mycobacterium spp. and to various adjuvants[J].J Fish Dis,1998,21(1):39-46.
[2] Dalmo R A,Seljelid R.The immunomodu1atory effect of LPS, laminaran and sulphated laminaran [β(1,3)-D-glucan] on Atlantic salmon, Salmo salar L.,macrophages in vitro[J].J Fish Dis,1995,18(2):175-185.
[3] 胡兵,刘军,侯永清,等.黄芪多糖对异育银鲫非特异性免疫力的影响[J].水利漁业,2008,28(3):108-111.
[4] 史会来, 楼宝, 胡则辉.中草药饲料添加剂对真鲷(Pagrosomus major)幼鱼生长和非特异性免疫的影响[J].现代生物医学进展,2007,7(4):548-551,556.
[5] 刘华忠,刘定忠,赵学明.复方中草药对彭泽鲫非特异性免疫功能的影响[J].淡水渔业,2004,34(3):31-32.
[6] 曹振杰,曲世科,丛日祥,等.免疫多糖对草鱼免疫功能的影响[J].齐鲁渔业,1999,16(3):43-44.
[7] Marta Alonso,Jo- Ann Leong.Suppressive subtraction libraries to identify interferon- inducible genes in fish[J].Marine Biotechnology,2002,4:74-80.
[8] Fletcher T C.Modulation of nonspecific host defense in fish[J].Vet Immunol I mmunopthol,1986,12(1-4):59-67.
[9] 韩璐,王彦波,王琼萍,等.细菌多糖对罗非鱼血清免疫功能影响的研究[J].饲料工业,2009,30(10):22-24.
[10] 汪成竹,姚鹃,吴凡,等.免疫多糖(酵母细胞壁)对中华鳖非特异性免疫功能的影响[J].华中农业大学学报,2006,8(6):421-425.
[11] 胡宗云,宋文华,富丽静,等.金丝桃素、绿原酸和硬葡聚糖对草鱼非特异性免疫功能的影响[J].水产学杂志,2014,27(1):41-45.
[12] 许国焕,吴月嫦,陶家发.两种多聚糖对彭泽鲫生长影响及免疫促进作用的初步研究[J].水利渔业,2002,22(4):49-51.
[13] 王树芹,周洪琪.壳聚糖对异育银鲫溶菌酶和白细胞吞噬活性的影响[J].上海水产大学学报,2004,13(2):121-125.
[14] 李桂峰,康裕财,孙际佳,等.酵母多糖对赤眼鳟非特异性免疫机能的影响[J].中山大学学报(自然科学版),2003(4):55-58.
[15] 李婵,徐奇友,许红,等.几种饲料添加剂对虹鳟(Oncrhynchus mykiss Walbaum)免疫活性和抗氧化能力的影响[J].安徽农业大学学报,2008,35(3):456-461.
[16] 周进, 黄倢, 宋晓玲.免疫增强剂在水产养殖中的应用[J].海洋水产研究,2003,24(4):70-79.
[17] 王洪斌,李士虎,阎斌伦.海洋红酵母多糖提取及其对日本蟳血清中部分免疫活性因子的影响[J].微生物学通报,2011, 38(2):176-180.
[18] 于翔,李赫,宋文华,等.三种免疫添加剂对草鱼非特异性免疫功能的影响[J].水产养殖,2011, 32(7):38-43.
[19] 刘丽萍,任翠爱,赵宏艳.甘草酸的免疫调节作用研究进展[J].中国实验方剂学杂志,2010, 16(6):272-276.
