

摘要:采用生长速度法研究了黑龙江省马铃薯黑痣病菌菌丝生长的条件、黑痣病菌的生物学特性及菌丝融合群情况,为病害的防治提供理论依据。结果表明,黑痣病病菌菌丝的适宜生长条件为25~30℃,低温情况下生长速度缓慢且在5℃不生长;最适合生长的pH值为7;黑暗环境更适合黑痣病病菌的生长;在本试验的几种碳源中,最利于病菌生长的是可溶性淀粉,在本试验的几种氮源中,最有利于病菌生长的是酵母粉。从黑龙江省7个马铃薯主产区采集马铃薯黑痣病样品100份,融合群测定结果表明,这些菌株分别属于AG-8、AG-6、AG-5、AG-4、AG-1-IC、AG-3和AG-1-IA融合群,其中AG-1-IA群有8个菌株,AG-1-IC群有13个菌株,AG-3群有10个菌株,AG-4群有20个菌株,AG-5群有17个菌株,AG-6群有10个菌株,AG-8群有14个菌株,其中AG-4为优势菌群。黑龙江省的马铃薯黑痣病菌融合群分布比较复杂,同一地点可能出现不同的融合群,同一融合群也会在不同的地区出现。
关键词:马铃薯;立枯丝核菌;生物学特性;菌丝融合群
马铃薯黑痣病是一种非常严重的土传病害,主要是由立枯丝核菌(Rhizoctonia solani)所引起,在马铃薯块茎及土壤中越冬存活。马铃薯黑痣病发病区域非常广泛,在世界各地均有发生。据报道,AG-3是立枯丝核菌主要的融合群,常常引起黑胫病、茎腐或匍匐茎腐烂,而AG-1、AG-2、AG-4、AG-5和AG-7等融合群也有相关研究报道。AG-2型病原菌存在3种基因型,其中AG-2-1550 bp的致病性是最强的,是导致匍匐茎和茎坏死的主要基因型,另外2个基因型的致病性较弱,仅会导致较小的病斑,而AG-3型病原菌主要对匍匐茎、块茎及茎进行侵染,致病能力很强。在田间试验和培养基中对块茎的检测都表明AG-3型病原菌其菌核的数量多于AG-2和AG-5型病原菌。AG-8型病原菌却只有对根部的侵染能力,对植株其他部位不具备侵染能力,而AG-7型病原菌却与之相反,不能侵染根部。生物学特性方面,李乾坤等对甘肃省马铃薯种植区采集的黑痣病菌菌株进行分离,研究表明,菌丝生长温度最低为4℃,最适为23℃,34℃时停止生长,刘宝玉等对内蒙古马铃薯种植区的马铃薯黑痣病菌菌株进行研究表明,菌丝生长最高pH为11,在pH为12时停止生长。
在黑龙江省,关于马铃薯黑痣病菌生物学特性的研究还比较少,融合群的鉴定分析研究接近空白,这使黑龙江省马铃薯黑痣病的防控缺少理论依据。笔者从黑龙江省7个马铃薯主产區采集分离得到100个马铃薯黑痣病菌菌株,通过对病菌生物学特性进行研究及菌丝融合群鉴定分析,为黑龙江省马铃薯黑痣病病害的发生及防治提供科学依据。
1材料与方法
1.1材料
2014-2015年从黑龙江克山、大庆、双城、绥化、克东、呼兰、黑河7个马铃薯种植区按种植面积的大小平均采集分离得到马铃薯黑痣病标本100份。标准菌株AG-8、AG-6、AG-5、AG-4、AG-1-IC、AG-3和AG-1-IA由黑龙江省农业科学院与中国农业大学植物保护学院提供。
1.2方法
1.2.1黑龙江马铃薯黑痣病菌生物学特性研究 从马铃薯块茎上选取菌核,先放入酒精中消毒30s,再在HgCl2中二次消毒2-3min,然后用无菌水冲洗,吸干表面水分后放人事先准备好的PDA上,保证每个PDA板上有3~4块菌核,培养温度设置在25℃,培养1~2d,待长出菌丝后,挑取尖端的菌丝,继续在PDA中纯化培养,判断立枯丝核菌的标准是有孢子、菌丝分枝或菌核的出现。确定是立枯丝核菌后,4℃保存在PDA斜面上备用。
将全部活化后的供试菌株转接到PDA平板上培养,待培养基长满菌丝后,用5mm的打孔器沿着培养基最外圈的菌丝进行打孔,然后将所得的所有菌碟转接到新的PDA培养基的正中心位置培养,处理36h后利用十字交叉法测量菌落的直径。
设置5、10、15、20、25、30、35℃共7个处理,3次重复,研究温度对菌丝生长的影响。
设置连续光照、连续黑暗、12h光暗交替3个处理,3次重复,研究光照对菌丝生长的影响。
得到菌碟后分别用1mol·L-1HCL和NaOH溶液调节培养基的pH,将菌碟分别放置在pH为3、4、5、6、7、8、9、10、11、12共10种培养基中,25℃恒温倒置培养,每个处理3次重复,研究pH对菌丝生长的影响。
查氏培养基:2g KNO3,1g KH2PO4,0.5g KCl,0.01g FeSO4,0.5g MgsO4·7H2O,20g琼脂,30g蔗糖,1000g蒸馏水。将以下4种碳源替代蔗糖,供试碳源:麦芽糖、可溶性淀粉、葡萄糖、果糖,以不加碳源作为对照;将以下4种氮源替代KNO3,供试氮源:酵母粉、尿素、蛋白质、硝酸钠,以不加氮源作为对照。分别设置5个处理,3次重复,将菌碟接种于不同碳源培养基中,25℃恒温培养,研究不同碳源、氮源对菌丝生长的影响。
1.2.2菌丝融合群鉴定 利用载玻片配对法将供试菌株和标准菌株的融合反应做鉴定试验,操作如下:首先,将事先准备好的标准菌株和待测菌株25℃恒温处理72h,然后利用打孔器选取5mm的菌落,在灭菌的载玻片上用移液枪打一层PDA培养基,将标准菌株放置在载玻片的中间,待测菌株放在标准菌株的两侧,间隔1.5-2.0cm,然后将该载玻片放入无菌培养皿加无菌水,25℃恒温培养24~36h,直到2种菌株边缘生长到一起并且交叉5mm左右时,利用显微镜观察菌丝融合的情况,3次重复。分3种情况进行鉴定,如表1所示。
2结果与分析
2.1黑龙江马铃薯黑痣病菌生物学特性
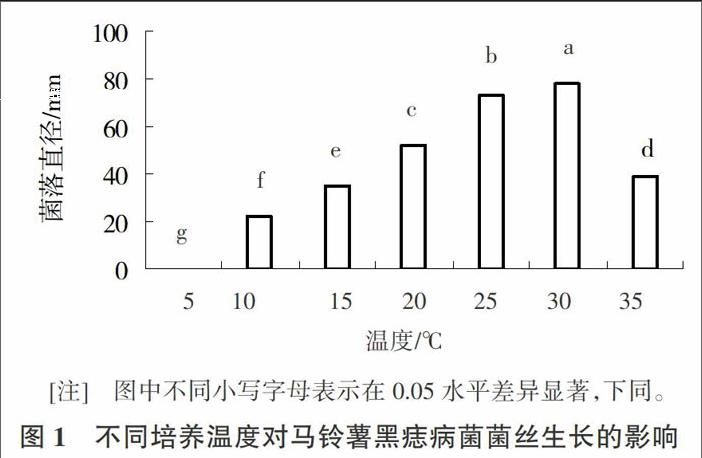
2.1.1温度对病菌菌丝生长的影响 病菌菌丝在不同培养温度下菌落生长速度不同,其中5℃条件下不生长、10~35℃条件下可以生长,且在10~30℃之间菌落直径随温度的升高明显增大,在30℃时达到最大值,当温度大于30℃时菌落直径随着温度的升高明显变小。因此,25~30℃的培养条件利于黑痣病菌生长,30℃是其最适培养温度。多重比较结果表明,菌株在不同温度处理间,菌絲生长速度差异显著(图1)。
2.1.2光照对菌丝生长的影响 对供试菌株病菌菌丝进行全光照、全黑暗、光暗交替处理,结果表明,全黑暗条件下菌丝生长速度最快,菌落平均直径为75.8mm;光暗交替条件下生长速度次之,菌落平均直径为71.8mm;在全光照条件下生长速度最慢;多重比较结果表明,全黑暗与光暗交替处理间差异不显著,但与全光照处理间差异显著(图2)。
2.1.3不同pH对菌丝生长的影响 病菌菌丝的生长在不同pH条件下速度不同,pH在3~12范围内马铃薯黑痣病菌菌丝均可生长,其中菌丝的生长速度较快的pH范围为5~8,且在pH=7时,菌丝生长速度最快,pH=12时,菌丝生长速度最慢。多重比较结果表明,pH=3和pH=4、pH=5和pH=8处理间的菌丝生长速度差异不显著,其余各pH处理间菌丝生长速度差异显著(图3)。
2.1.4不同碳源对菌丝生长的影响 由表2可知,病菌在供试的5种碳源中均可生长,其中,在可溶性淀粉培养基中生长的菌落直径最大,蔗糖其次,而在果糖培养基中生长的菌落直径最小。
2.1.5不同氮源对菌丝生长的影响 由表3可知,病菌在所提供的5种氮源中均可生长,其中,在酵母粉培养基中生长的菌落直径最大,蛋白胨培养基其次,而在尿素培养基中菌丝生长效果最差。
2.2黑龙江马铃薯黑痣病菌融合群鉴定
2.2.1菌丝融合观察 对菌丝融合进行观察是很直观的检测融合结果的方法,但菌丝融合存在不同的形式,因此,根据不同的融合形态将菌丝融合分为3种类型:A-不融合,B-完全融合,C-不完全融合(图4)。其中完全融合也分为3种情况,顶端对顶端的融合、顶端对侧枝的融合、侧枝对侧枝的融合。
2.2.2菌丝融合群的鉴定 对所收集的黑龙江省双城(SC)、大庆(DQ)、克山(KS)、克东(KD)、黑河(HH)、呼兰(HL)、绥化(sH)共7个地区的病菌菌株进行菌丝融合群鉴定,根据7个标准菌株可将供试菌株分为7个群,其中,AG-1-IA群有8个菌株,AG-1-IC群有13个菌株,AG-3群有10个菌株,AG-4群有20个菌株,AG-5群有17个菌株,AG-6群有10个菌株,AG-8群有14个菌株(表4)。说明马铃薯立枯丝核菌菌丝融合群在黑龙江省的分布是较广泛的。由以上数据可知,AG-4的分离频率最大,其次是AG-5,AG-8,AG-I-IC,AG-3,AG-6和AG-I-IA。
2.2.3马铃薯黑痣病菌融合群分布频率 对收集的100个菌株经过融合群分析鉴定之后,将黑龙江省的菌株分为7个群,分别是AG-4、AG-5、AG-6、AG-8、AG-1-IC、AG-3和AG-1-IA。其中,AG-I-IA在克山县、绥化市、大庆市和呼兰区均有分布,AG-I-IC在大庆市、克山县、克东县和双城区均有分布,AG-3在双城区、克山县和绥化市有分布,AG-4在绥化、大庆市、克山县、双城区均有分布,AG-5在双城区、绥化市、大庆市、呼兰区、克山县和黑河均有分布,AG-6在双城区、大庆市、绥化市和呼兰区均有分布,AG-8在克东县、绥化市、克山县、大庆市均有分布。其中AG-4出现频率最高,为20%,是优势菌群(表5)。
3讨论与结论
马铃薯黑痣病在我国最早是1922年在台湾发现的,到目前为止,它的发生已经非常普遍。病菌侵染幼芽,导致幼芽出土前腐烂甚至死亡形成芽腐,造成缺苗;侵染植株茎基产生凹陷褐色病斑,严重的导致立枯、叶片卷曲及顶端萎蔫的症状;侵染薯块导致其表面产生大小、形状不规则的黑色菌核。本研究结果表明,马铃薯黑痣病菌菌丝适宜生长条件为25~30℃,低温情况下生长速度缓慢且在5℃以下不生长;而甘肃省马铃薯种植区采集的黑痣病菌菌株菌丝生长温度最低为4℃,最适为23℃,34℃时停止生长。笔者分离菌株的菌丝最适生长pH为7;而内蒙古马铃薯种植区的马铃薯黑痣病菌菌丝最高生长pH为11,在pH为12时则停止生长,并且黑暗环境更适合病菌生长。笔者比较了几种不同碳源和氮源对马铃薯黑痣病菌菌丝生长的影响,结果表明,在几种碳源中,最利于病菌生长的是可溶性淀粉,而在参与试验的几种氮源中,最有利于病菌生长的是酵母粉,这为以后黑痣病菌的室内培养和研究提供了参考资料。对比国内其他研究结果和本研究结果可知,马铃薯黑痣病菌随地域的不同而具有不同生长特性,并且适应特定的生长环境,其原因可能是病原菌属于不同群属,且在当地特定环境中不断进化适应。试验得出的黑龙江马铃薯黑痣病菌适宜生长条件为黑龙江省马铃薯黑痣病的防治提供了理论依据,在实际应用中,在病菌适宜生长条件出现时能够及时地进行预防。
有关立枯丝核菌分类,前人已经有相关研究,目前被广泛认可的分类方法是以菌丝融合的方式来确定融合群。利用菌丝融合群的方法,前人进行了亲缘关系的研究。这些研究结果表明,融合群之间的差异明显大于融合群内独立菌株的差异。因此,菌丝融合群的分类能够鉴别出不同类型的种群。立枯丝核菌主要来源于水稻、小麦和棉花等,能够引起多种植物发生病害,且其融合群较为多样。例如AG-4是新疆和四川棉花上的主要融合群;AG-1-IA是四川、江苏和广西等地水稻和玉米上的优势融合群;AG-1和AG-2是上海、浙江、山东、河南以及陕西草坪上检测到的立枯丝核菌融合群;AG-5是湖北小麦上立枯病丝核菌主要融合群。本试验将黑龙江省马铃薯黑痣病融合群主要分为AG-8、AG-6、AG-5、AG-4、AG-I-IC、AG-3和AG-1-IA,其中AG-1-IA群有8个菌株,AG-1-IC群有13个菌株,AG-3群有10个菌株,AG-4群有20个菌株,AG-5群有17个菌株,AG-6群有10个菌株,AG-8群有14个菌株。另外有8个菌株没有检测到融合群,这8个菌株与所选标准菌株均不融合,其原因可能是本试验所选用的标准菌株只有7株,并不能排除黑龙江省马铃薯黑痣病菌可能存在其他融合群,还有待于进一步研究。笔者还对黑龙江省马铃薯黑痣病菌菌丝融合群的分布进行了统计,结果表明,其分布非常广泛,在分布上存在地域差异性。并且有别于中国北方马铃薯黑痣病菌融合群鉴定及遗传多样性分析试验中AG-3是优势菌群、AG-4是次优势菌群的结果,本研究结果表明,AG-4为主要融合群,AG-3所占比例很少,此差异可能是所选取的菌株分离地域差异和试验材料的生态类型差异所引起。对不同地域优势菌群的研究有利于对病原菌进行针对性防治,为缓解该病原菌引起的病害和减少病原菌引起的农业生产损失提供宝贵的理论支持。
- 论电信企业营销渠道管理系统的设计与应用
- 重资产企业供应链管理优化的思考
- 构建电网施工企业内控体系 提升国际业务风险防控水平
- 区域市场营销与企业市场营销的关系研究
- 新形势下国有企业成本管理问题研究
- 浅谈变电运行的安全管理和故障排除
- 浅谈假发票的鉴别及防治机制的建设
- 浅谈新常态下做好实物保护工作的体会和建议
- 政府科研经费管理与效益研究
- 房产管理的实践与探索
- 试析路政管理对促进经济发展的影响
- 我国社会保险基金的内部控制问题探究
- 浅谈“营改增”对煤炭企业税收筹划的影响及对策
- 企业经济效益审计风险规避及策略选择
- 基于“营改增”下的小区配电工程税收筹划
- 建筑施工企业税务风险管理研究
- 浅谈如何开展“两学一做”教育活动做合格党员
- 市场经济形势下党员干部更要注重学习党章,提高党性修养
- 关于新形势下提升企业基层支部战斗力的探索
- 离退休人员的思想特点、难点和解决策略分析
- 论新常态下求真务实创新工会工作
- 企业的合并和发展
- 城市记忆档案的特点及开发利用
- 失地农民市民化、后续管理与产业发展
- 锅炉安装和调试中的重点技术探讨
- cash issue
- cashissue
- cashless
- cashmachine
- cash machine
- cash machines'
- cashmanagement
- cash management
- cashmarket
- cash market
- cash maˌchine
- cashmere
- cashmeres
- cashmerian
- cashmerians
- cash out
- cash pile
- cashpile
- cashpoint
- cashpoint™
- cashposition
- cash poˌsition
- cashprice
- cash price
- cashratio
- long
- lonɡ
- lou
- lpc
- lq
- lq
- ltns
- ltv
- ltv
- lu
- luan
- lue
- lun
- luo
- luàn
- luán
- luò
- luó
- luō
- luǎn
- luǒ
- luɑn
- là
- lài
- làn