蔡燕丽 刘欢 王翀
摘要:对新疆石河子和吐鲁番2个地区酿酒葡萄园可培养酵母菌的种类鉴定及分布进行研究,旨在明确酿酒葡萄园主要酵母种类及分布规律,为酵母菌资源的开发利用打下基础。对酿酒葡萄园果实表面和土壤进行稀释后平板划线分离纯化,挑取单菌落,并对菌落和细胞显微特征进行形态观察。对分离获得的酵母菌株进行WL培养基培养观察,区分聚类,选取其代表菌株进行糖发酵及碳源利用试验,且对其26S rDNA D1/D2区序列测序鉴定并进行系统进化分析。结果发现,从酿酒葡萄园土壤及果实表面中共分离获得41株酵母菌株,其中在石河子地区分离获得28株,经鉴定属于隐球酵母属(Cryptococcus)、有孢汉逊酵母属(Hanseniaspora)、威克汉姆酵母属(Wickerhamomyces)、毕赤酵母属(Pichia)、棒孢酵母属(Clavispora)、红冬孢酵母属(Rhodosporidium)、红酵母属(Rhodotorula)、假丝酵母属(Candida)、Rhodosporidiobolus和Kazachstania等10属13种,在吐鲁番地区共分离获得13株,经鉴定属于棒孢酵母属(Clavispora)、假丝酵母属(Candida)、红酵母属(Rhodotorula)、隐球酵母属(Cryptococcus)、Kazachstania和Rhodosporidiobolus等6属6种。结果表明,新疆酿酒葡萄园的酵母菌组成具有较为丰富的多样性,其应用价值值得进一步研究。
关键词:新疆酿酒葡萄;酵母菌;多样性分析;26S rDNA
中图分类号: S182 ?文献标志码: A ?文章编号:1002-1302(2019)02-0282-05
酿酒葡萄首要用途就是用来酿造葡萄酒。2014年新疆葡萄种植面积占全国葡萄面积的19.4%,产量占全国总产量的18.5%,居全国首位,而且产量持续增长[1]。新疆因其有特殊的气候和生境而适宜葡萄的种植。新疆酿酒葡萄的种植主要集中在吐鲁番、石河子周边地区。近年,国内对于酿酒葡萄的研究主要集中于地域性生境对酿酒葡萄种植的影响[2]和微生物群落多样性分析[3],少有对酿酒葡萄园中酵母菌的研究。新疆日照充沛,光合效率高,糖分积累容易,为酵母菌提供了适宜的生长环境,同时新疆昼夜温差大的自然条件为酵母菌适应性进化提供了驱动力[4],因此新疆酵母菌多样性具有特殊性。
本研究采样地点主要集中在吐鲁番和石河子地区,它们分别位于41°12′~43°40′N,87°1′″~91°55′E之间和43°26′~45°20′N,84°58′~86°24′E之间,属于独特的暖温带大陆性干旱荒漠气候和典型的温带大陆性高原气候。本研究以酿酒葡萄园采集的土壤和果實为原料,在适宜的培养条件下分离筛选其中的酵母菌菌株,并进行传统的形态分类和分子生物学鉴定,探明新疆酿酒葡萄主产区可培养酵母菌菌群组成和分布。
1 材料与方法
1.1 材料与试剂
1.1.1 样品采集 2016年8月在吐鲁番与石河子地区分别选取1个酿酒葡萄园,采集样地属于规模化酿酒葡萄园,葡萄树龄在3~5年。分别在2个酿酒葡萄园中采集土壤和果实样品,土壤按“S”形5点取样混合法取样,采集好的土壤和果实样品装入灭菌信封,放入冰盒,带回石河子大学实验室,放入4 ℃冰箱中保存,用于酵母菌的分离鉴定。
1.1.2 培养基[5]
1.1.2.1 YPD液体培养基 酵母浸粉10 g/L,蛋白胨 20 g/L,葡萄糖20 g/L。
1.1.2.2 PDA培养基 马铃薯200 g/L,葡萄糖20 g/L,琼脂粉20 g/L,pH值为6.0。
1.1.2.3 WL琼脂培养 酵母浸粉4 g/L,胰蛋白胨5 g/L,葡萄糖50 g/L,磷酸二氢钾550 mg/L,氯化钾425 mg/L,氯化钙125 mg/L,硫酸镁125 mg/L,氯化铁2.5 mg/L,硫酸锰 2.5 mg/L,溴甲酚绿22 mg/L,琼脂 20 g/L,pH值为5.5。
1.1.2.4 糖类发酵培养基 6 g/L酵母粉,20 g/L 糖,8磅 15 min 灭菌。发酵用糖类包括葡萄糖、乳糖、半乳糖、麦芽糖、蜜二糖、蔗糖和棉子糖等。
1.1.2.5 碳源同化基础培养基 (NH4)2SO4 5 g/L,KH2PO4 1 g/L,MgSO4·7H2O 0.5 g/L,CaCl2·2H2O 0.1 g/L,NaCl 0.1 g/L,酵母粉 0.2 g/L,碳源5 g/L,8磅15 min灭菌。同化试验用碳源包括葡萄糖、乳糖、半乳糖、麦芽糖、蜜二糖、蔗糖、棉子糖、纤维二糖、海藻糖、D-阿拉伯糖、L-阿拉伯糖和可溶性淀粉等。
1.2 仪器与设备
ZHWY-100B摇床(上海智城分析仪器制造有限公司);HWS恒温恒湿培养箱(上海精宏实验设备有限公司);FroFleX PCR仪(美国应用生物系统公司);5424R高速冷冻离心机(Eppendorf公司);SW-CJ-1F超净工作台(苏州净化设备有限公司)。
1.3 方法
1.3.1 菌株分离与保藏[5] 称取5 g分离样品,倒入装有 50 mL 灭菌YPD液体培养基的锥形瓶中,摇培24~48 h。然后在超净工作台上按10倍稀释法进行稀释分离。移取稀释液0.1 mL涂布于PDA固体培养基上,28 ℃恒温培养48~72 h。挑取单菌落在PDA固体培养基上近一步划线纯化2~3次,获得纯培养物。纯化菌株采用PDA斜面和干粉冷冻保藏[6-7]。
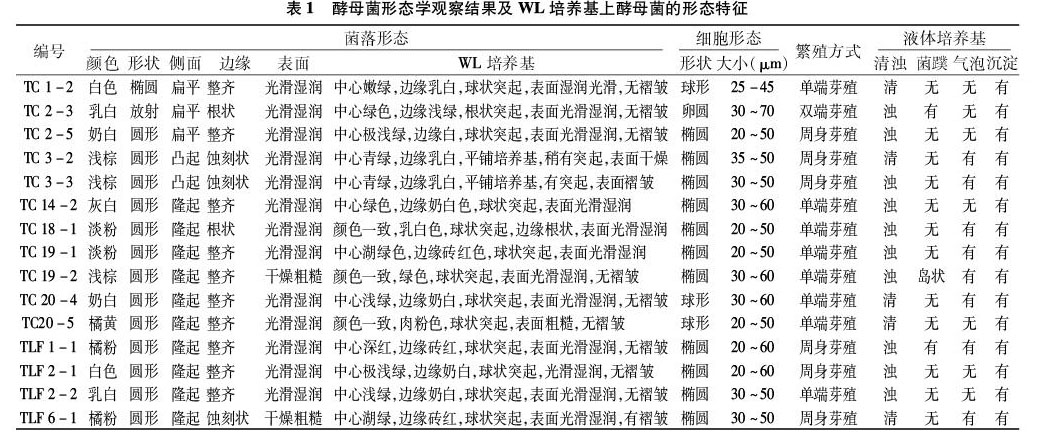
1.3.2 酵母菌形态聚类分析 根据酵母菌分类学的标准研究方法,对菌株进行菌落形态和细胞显微形态分析[5]。同时将单菌落划线于WL鉴别培养基上28 ℃恒温培养3~4 d[8]。依据菌落特征和细胞颜色等不同对分离菌株进行初步归类。菌落特征包括菌落形态、色泽、湿润程度、表面是否光滑、边缘是否整齐等。细胞显微形态包括细胞形状、大小、繁殖方式、是否有假菌丝等。
1.3.3 酵母菌生理生化指标 对分离获得的菌株进行生理生化鉴定[5,9]。
1.3.4 酵母菌26S rDNA D1/D2区序列分析及系统发育分析 将获得的酵母细胞接种在5 mL YPD液体培养基中 28 ℃ 培养过夜,离心收集细胞,用CTAB法提取DNA后进行26S rRNA基因的PCR扩增。PCR扩增体系:DNA 1 μL、10×Taq buffer 2.5 μL、25 mmol/L MgCl2 2.5 μL、10 mmol dNTP 2 μL,NL-1 1 μL,NL-4 1 μL、Taq DNA聚合酶0.5 μL,ddH2O定容至总体积25 μL。PCR程序为94 ℃ 5 min;94 ℃ 45 s,56 ℃ 45 s,72 ℃ 1 min 30 s,30个循环;72 ℃ 10 min,4 ℃ 保存[6]。26S rRNA基因的扩增引物为NL-1:5′-GCATATCAATAAGCGGAGGAAAAG-3′,NL-4:5′-GGTCC GTGTTTCAAGACGG-3′。将扩增出的26S rRNA基因序列克隆送生工生物工程(上海)股份有限公司测序。登录NCBI网站(http://www.ncbi.nlm.nih.gov/)用BLAST程序进行序列比对,下载序列,通过Mega软件中的邻接法(neighbor-Joining)进行分析构建进化树。
2 结果与分析
2.1 酵母菌形态观察及聚类分析
经分离纯化,从样品中共分离出41株酵母菌,采用传统的形态学方法观察以及WL鉴别培养基培养,依据在WL培养基上长出的酵母菌菌落形态和颜色特征,结合在PDA上生长的酵母菌菌落及菌体形态可初步将分离菌株分为15种形态类型(表1、图1)。WL鉴别培养基生长鉴定结果见表2。初步分析显示,石河子地区分离酵母菌形态类型较吐鲁番地区多样。
2.2 酵母菌生理生化分析
对经初步筛选过的酵母菌分离株进行糖发酵、碳同化以及温度耐受性试验(表2、表3),可以看出,不同的酵母菌株类型对不同碳源的利用各不相同,同时进一步显示了新疆不同地区酵母菌多样性之间的差异和潜在的酵母菌资源。
2.3 酵母菌株26S rDNA测序分析
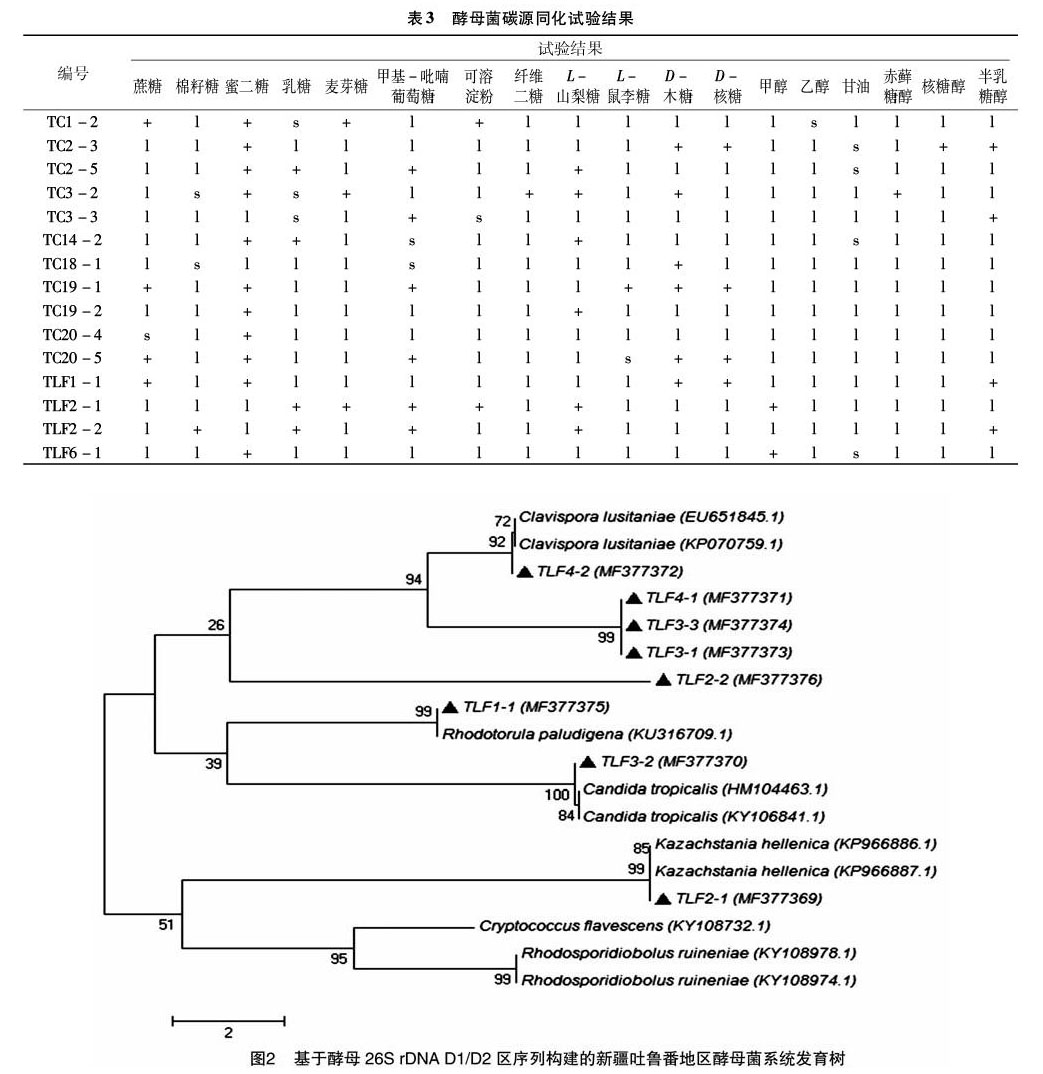
本研究在进行酵母菌形态聚类分析和生理生化分析的基础上,从每种形态类型中选择1株酵母菌进行26S rDNA基因扩增,PCR扩增产物均在550~700 bp之间。然后将PCR产物送生工生物工程(上海)股份有限公司进行测序,将测序所得的序列在GenBank上进行Blast比对,选取下载1~2个相似度较高菌株的26S rDNA序列,利用MEGA构建系统发育树(图2、图3)。系统发育树结果显示,各供试菌株与各自的模式菌株聚在一枝,并且相同种的菌株归于同一分枝,具有较近的亲缘关系,而不同种则位于不同分枝上。系统发育树的聚类结果与26S rDNA D1/D2区碱基序列的同源率鉴定结果相一致。
2.4 不同地区酵母菌种群分布及优势类群
本研究在石河子和吐鲁番酿酒葡萄园区分别采集葡萄样品和土壤样品,筛选出酵母菌株41株。经形态学鉴定,初步分为15类,选取分别来源于2个不同地区的代表性菌株28株, 对其进行DNA提取及PCR扩增、 26S rDNA D1/D2区域序列鉴定,结果显示,分离菌株属于隐球酵母属(Cryptococcus)、汉逊酵母属(Hanseniaspora)、威克汉姆酵母属(Wickerhamomyces)、毕赤酵母属(Pichia)、棒孢酵母属(Clavispora)、紅酵母属(Rhodotorula)、假丝酵母属(Candida)、Rhodosporidiobolus和Kazachstania,共9个属15个种。石河子地区分离获得酵母菌株28株,分别属于乌兹别克斯坦隐球酵母(Cryptococcus uzbekistanensis)、浅白隐球酵母(Cryptococcus albidus)、葡萄汁有孢汉逊酵母(Hanseniaspora uvarum)、异常威克汉姆酵母(Wickerhamomyces anomalus)、库德毕赤酵母(Pichia kudriavzevii)、葡萄牙棒孢酵母(Clavispora lusitaniae)、红冬孢酵母(Rhodosporidium kratochvilovae)、红冬孢酵母(Rhodosporidium babjevae)、热带假丝酵母(Candida tropicalis)、毕赤膜酵母(Pichia membranifaciens)、浅黄隐球酵母(Cryptococcus flavescens)、Kazachstania hellenica和Rhodotorula nothofagi等10属13种,优势酵母菌种为异常威克汉姆酵母(Wickerhamomyces anomalus),占分离菌株的 17.9%。吐鲁番地区共分离获得13株酵母菌,经鉴定属于葡萄牙棒孢酵母(Clavispora lusitaniae)、热带假丝酵母(Candida tropicalis)、浅黄隐球酵母(Cryptococcus flavescens)、沼泽生红冬孢酵母(Rhodosporidium paludigena)、Rhodosporidiobolus ruineniae和Kazachstania hellenica等6属6种,优势酵母菌种为葡萄牙棒孢酵母(Clavispora lusitaniae),占分离菌株的46.2%。
3 结论与讨论
新疆地理环境特殊,不同地区有其独特的生态环境,同时酿酒葡萄种植面积广,不同产区的生态环境差异较大,因而酵母菌资源非常丰富[10]。本研究从2个酿酒葡萄园中共分离41株酵母菌,分布于9个属15个种,分别为乌兹别克斯坦隐球酵母(Cryptococcus uzbekistanensis)、浅白隐球酵母(Cryptococcus albidus)、葡萄汁有孢汉逊酵母(Hanseniaspora uvarum)、 异常威克汉姆酵母(Wickerhamomyces anomalus)、库德毕赤酵母(Pichia kudriavzevii)、葡萄牙棒孢酵母(Clavispora lusitaniae)、红冬孢酵母(Rhodosporidium kratochvilovae)、红冬孢酵母(Rhodosporidium babjevae)、热带假丝酵母(Candida tropicalis)、毕赤膜酵母(Pichia membranifaciens)、浅黄隐球酵母(Cryptococcus flavescens)、沼泽生红冬孢酵母(Rhodosporidium paludigena)、Rhodotorula nothofagi、Kazachstania hellenica、Rhodosporidiobolus ruineniae,其中Clavispora lusitaniae、Kazachstania hellenica、Candida tropicalis和Cryptococcus flavescens在2个酿酒葡萄园中均有分布。此外,本研究结果显示石河子葡萄园中分离的可培养酵母菌比吐鲁番地区可培养酵母菌多样性大。李双石等研究山东蓬莱和甘肃武威的蛇龙珠葡萄附着酵母菌的种群多样性及其生态分布特征发现,不同葡萄产区由于气候、土壤和管理方式等不同,即使是同一葡萄品种分布的酵母菌种也存在差异[11],本研究结果与之一致。本研究发现石河子酿酒葡萄园酵母菌优势菌种为异常威克汉姆酵母(Wickerhamomyces anomalus),占分离菌株的17.9%。吐鲁番酵母菌优势菌种为葡萄牙棒孢酵母(Clavispora lusitaniae),占分离菌株的46.2%。这与李艳等对沙城地区3个龙眼葡萄园酵母菌的研究结果[12]相一致,进一步说明不同来源的土壤样品中酵母菌群组成各不相同,优势菌种的类型也不相同。
- 基于价值链理论的企业成本管理模式
- 基于风险导向的工程建设企业内控体系的构建
- 集团内部控制问题研究和对策分析
- 试析风险管理视角下高新技术企业内控制度的实施与应用
- 推进邮政网点损益核算的策略探析
- 浅析物业企业成本管理
- 浅析集团化旅游公司中内部控制体系建设的重要性
- 对建立企业内部管理控制系统框架的探讨
- 产业新城PPP项目股权合作合并报表问题的探讨
- 基于连锁餐饮企业的内部控制制度建设研究
- 商业银行个人信贷业务的潜在风险及其处理对策分析
- 以全面预算管理为基础的企业内部控制研究
- 分析企业现金流量管理与风险防范
- 发电企业的资金预算与风险防控分析
- 科学事业单位科研经费管理存在的问题及优化建议
- 商业银行互联网信贷业务的风险管理研究
- 谈《资产评估法》实施后的行业行政监管
- 试论施工企业如何有效加强现金流管理及控制
- 成本精细化管理在石油企业中的应用浅析
- 公司存货管理内部控制分析
- 深化燃煤发电上网电价形成机制改革答记者问
- 关于煤炭火力发电企业内审实务的探讨
- 公立医院内部审计信息化建设的实践与探索
- 内部审计在医院经济管理中的重要性分析
- 浅谈路桥施工企业内部审计存在的不足与解决措施
- proper-nouns'
- propers
- properties
- property
- propertybond
- property bond
- property company
- propertycompany
- propertydevelopment
- property deˌveloper
- property deˌvelopment
- propertyless
- propertylesses
- propertylessness
- propertylessnesses
- propertyloan
- property loan
- property management
- propertymanagement
- property manager
- property market
- propertymarket
- property register
- propertyregister
- property tax
- 春色满眼
- 春色满眼弥望
- 春色美丽
- 春色美好
- 春色美好,风光无限
- 春色迷人
- 春艳
- 春节
- 春节包机
- 春节吃什么
- 春节妈妈
- 春节期间
- 春节期间的饮宴
- 春节用红纸写好贴在门上的联
- 春芜
- 春花
- 春花然然
- 春花秋实秋实春华
- 春花秋月
- 春芳
- 春芽
- 春苗
- 春苗得雨——正逢时
- 春苗茁壮
- 春英