孔瑞 胡正华 陈书涛 柯浩楠 王亚萍 吴杨周
摘要:通过田间控制试验,设置对照(CK)、增温约2 ℃(T)、降水量减少30%(P)、增温约2 ℃+降水量减少30%(TP)4种不同处理,研究增温和降水减少对作物生物量以及叶片硝酸还原酶(nitrate reductase,简称NR)、土壤酶活性的影响。结果表明,在冬小麦生长季,与CK相比,T处理在抽穗—扬花期显著增加地上生物量46.01%(P=0.000)、总生物量34.07%(P=0.001),在灌浆—成熟期显著增加地上生物量30.47%(P=0.028);TP处理在拔节—孕穗期显著增加地上生物量 53.45%(P=0.024),在抽穗—扬花期显著增加地上生物量19.82%(P=0.014),在收获期显著增加地上生物量 34.60%(P=0.004)、总生物量29.08%(P=0.011)。在大豆生长季,与CK相比,T处理在三叶期显著增加地下生物量48.89%(P=0.035)、总生物量38.23%(P=0.041),在开花期显著增加地上生物量82.61%(P=0.035)、总生物量74.05%(P=0.040);TP处理在三叶期显著增加地上生物量43.56%(P=0.030)、总生物量4254%(P=0.027);P处理对冬小麦和大豆的生物量影响未达到显著水平。增温、降水量减少处理对冬小麦和大豆叶片NR活性没有显著影响,但TP处理在大豆开花期显著增加NR活性,增幅为42.84%(P=0.023)。增温和降水量减少对冬小麦和大豆土壤酶活性的影响规律基本一致,相比于CK,T处理增加土壤转化酶和脲酶活性、降低土壤过氧化氢酶活性,P处理降低扬花期土壤脲酶活性,TP处理增加成熟期土壤转化酶活性。
关键词:增温;降水量减少;冬小麦;大豆;生物量;酶活性
中图分类号: S181;S162.5 文献标志码: A 文章编号:1002-1302(2019)23-0110-06
21世纪全球的平均气温将会升高1.5~4.8 ℃[1],全球气温升高且具有持续上升的趋势已成为不争的事实。地表温度的持续升高将会改变大气环流及水文模式,进而引发全球或区域降水格局的变化[2-3],极端降水和干旱事件随之交替频发[4]。降水格局和温度作为影响农田生态系统植物生长和土壤微生物活性的重要因素,由气候变化引起的温度升高和降水格局变化势必会对农田生态系统产生影响。
农田在陆地生态系统的物质循环和能量流动中起着主导作用,农作物生物量是研究全球碳循环的重要组成部分,作为评价生态系统的重要指标,许多学者对不同条件下农作物生物量对温度和降水量等环境因子改变的响应进行研究[5-10]。植物叶片中的硝酸还原酶(nitrate reductase,简称NR)为植物体内尿素同化和代谢过程中的关键限速酶,可反映植株的氮素营养状况和氮代谢水平[10-11];脲酶可表征土壤氮素状况,其活性与土壤的微生物数量、有机物质含量呈正相关关系;转化酶水解蔗糖为葡萄糖和果糖,其活性可表征土壤肥力与熟化程度;过氧化氢酶分解由生物呼吸和有机物氧化产生的过氧化氢,其活性在一定程度上可表征土壤总的生物学活性和肥力状况[12]。已有研究表明,增温不仅降低植物叶绿素含量和光合速率[13],缩短作物生育期[14-15],还会加速土壤有机质分解,增加土壤微生物生物量和活性[16],土壤酶活性也随之改变。作为影响农田生物量和酶活性的关键因素,土壤条件和作物生理生长状况受降水量变化影响较大[17],因此,研究增温和降水量减少对作物生物量和酶活性影响有重要意义。
目前,国内外关于增温和降水量变化对作物生物量复合效应影响的研究在农田生态系统中已取得部分进展[9,18-20],针对酶活性的研究却多见于单因子影响[16-17,21-23],而针对不同农作物品种的对比研究较少。本研究利用红外辐射增温装置模拟田间增温,人工控制水分变化,通过测定冬小麦和大豆不同生育期生物量、叶片NR活性和3种土壤酶活性,探究增温和降水量减少的协同作用对不同农作物生物量和酶活性的影响规律,以期為准确评价在全球气候变化背景下,我国华东地区农田生态系统作物生长和土壤微生物对生态系统环境变化的响应规律与机制提供基础性资料,为国内粮食安全保障研究提供理论支撑。
1 材料与方法
1.1 田间概况
本研究于2013—2014年冬小麦和大豆生长季在南京信息工程大学农业气象试验站(32.21°N,118.71°E)进行大田试验。该地多年平均降水量为1 100 mm,平均温度为 15.6 ℃。试验地土壤为潴育型水稻土,灰马干土属,耕层土壤质地为壤质黏土,黏粒含量为26.1%,土壤pH(H2O)值为6.64,有机碳、全氮含量分别为35.49、0.55 g/kg。
1.2 试验方法
1.2.1 试验设计 试验共设置4种处理,分别为对照(CK)、增温约2 ℃(T,比CK叶面温度增加约2 ℃)[21]、降水量减少30%(P)[9]、增温约2 ℃+降水量减少30%(TP)。每种处理分别设3个重复试验小区,共12个小区,各小区随机排列。每个小区面积为2 m×2 m,为防止小区之间的物质交换,相邻小区之间间隔0.5 m,并挖40 cm深的排水沟。模拟增温与降水量减少处理的时间为作物出苗到收获。
供试作物品种为当地常规种植的冬小麦(扬麦16)和大豆(八月白),冬小麦和大豆的主要生育期和肥料管理见表1。
采用红外线辐射器(infrared radiators)增温装置对土壤-作物系统进行昼夜不间断增温处理。每个增温小区设3个灯罩,灯罩内安装红外线辐射灯管(长1 m,功率250 W),平行悬挂在已安装好的不锈钢支架上,保持其高度在作物冠层上方1 m处。不作增温处理的小区,悬挂相同不通电灯罩,以降低或消除光照、遮阴等因素的干扰。所有灯罩高度都随作物生长高度进行调节。这种增温方法能较好地模拟全球变暖机制,且安装简单、维护方便,是国内外学者常用的较为理想的增温方式[21,24]。
降水量减少处理采用人工遮挡的方法,在P、TP处理试验小区上方均匀搭建6块透明聚氯乙烯(PVC)材料的阳光板作为遮雨板。每块遮雨板长×宽为2.5 m×0.1 m,距离地面高度为1.5 m,总截面积为试验小区面积的30%,以此达到减少30%降水的效果。遮雨板横向凹曲呈“U”形,与水平面保持一定坡度悬挂,便于降水顺着遮雨板流下进入导水沟渠。这种降水量减少的方法便于操作,在实际研究中得到广泛应用,如程徐冰等利用人工截留降水的方法研究长白山蒙古栎叶片生理生态特征对降水量减少的响应[25]。
1.2.2 样品采集与相关指标测定 分别在冬小麦关键生育期(返青、拔节、孕穗、抽穗、灌浆期)、大豆关键生育期(分枝、开花、结荚、鼓粒、成熟期),避开降水、施肥期,采用常规方法测定植株生物量以及土壤脲酶、转化酶、过氧化氢酶活性,其中植株叶片的硝酸还原酶活性在小麦关键生育期(扬花、灌浆、成熟期)、大豆关键生育期(开花、结荚期)测定。
(1)叶片硝酸还原酶活性的测定:在每个小区随机剪取长势均匀的作物叶片若干,用磺胺比色法测定叶片硝酸还原酶活性[11]。
(2)作物生物量测定:在每个小区随机选取1株长势均匀的作物植株连根系及周围土壤一同挖出,洗净后分地上和地下部分,置于恒温箱中在105 ℃下杀青0.5 h,然后在80 ℃下烘干至恒质量,分别称量地上、地下部分质量。
(3)土壤脲酶活性测定:取100 g左右根际土壤,风干,研磨后过20目筛,用靛酚比色法测定土壤脲酶活性[11]。
(4)土壤转化酶活性测定:取2 g过20目筛的风干土,用比色法测定葡萄糖生成量表示土壤转化酶酶活性[11]。
(5)土壤过氧化氢酶活性测定:在关键生育期,避开降水和施肥期,取5 g过1.25 mm筛的风干土,用比色法测定葡萄糖生成量,表示土壤过氧化氢酶活性[11]。
1.3 统计分析
采用Microsoft Office Excel 2010进行数据分析,数据结果用平均值±标准误差来表示;运用SPSS 19.0(SPSS Inc.,Chicago,USA)进行统计分析,采用LSD法进行不同处理间的差异显著性检验,运用OriginPro 2016(OriginPro,USA)软件完成制图。
2 结果与分析
2.1 增温和降水量减少对生物量的影响
2.1.1 冬小麦生物量 与CK相比,T处理地上生物量在抽穗—扬花期极显著增加46.01%(P=0.000)、灌浆—成熟期显著增加30.47%(P=0.028);TP处理地上生物量在拔节—孕穗期显著增加53.45%(P=0.024)、抽穗—扬花期显著增加19.82%(P=0.014),收获时地上生物量达到极显著增加的水平,增幅为34.60%(P=0.004);P处理对冬小麦地上生物量无显著影响(图1-a)。
与CK相比,T、P、TP处理对地下生物量没有显著影响。但从整个生育期来看,T处理呈现减少地下生物量的趋势,收获时,TP、P处理使地下生物量呈减少趋势,但未达到显著水平(图1-b)。
与CK相比,T处理总生物量在拔节—孕穗期显著增加34.07%(P=0.001);TP处理总生物量在收获时显著增加29.08%(P=0.011);P处理对冬小麦总生物量无明显影响(图1-c)。
2.1.2 大豆生物量 与CK相比,T处理下开花期大豆地上生物量显著增加82.61%(P=0.035);TP处理下三叶期大豆地上生物量显著增加43.56%(P=0.030);P处理对大豆地上生物量无显著影响(图2-a)。
与CK相比,T处理下三叶期大豆地下生物量显著增加48.89%(P=0.035),其他生育期无显著影响;P、TP处理对大豆地下生物量无显著影响,但P处理下大豆地下生物量整體呈现减少的趋势(图1-b)。
与CK相比,T处理下总生物量在三叶期显著增加 38.23%(P=0.041)、开花期显著增加74.05%(P=0.040);TP处理下,总生物量在三叶期显著增加42.54%(P=0.027);P处理对大豆总生物量无显著影响(图2-c)。
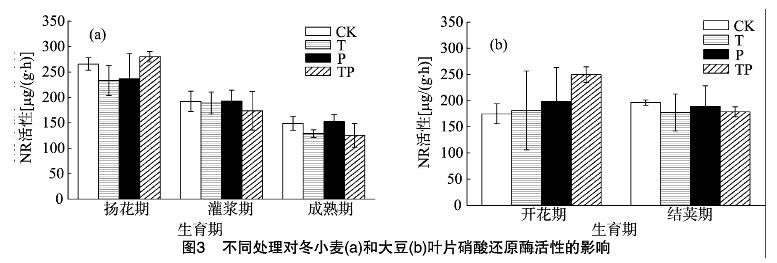
2.2 增温和降水量减少对叶片NR活性的影响
2.2.1 冬小麦叶片NR活性 与CK相比,3种处理对冬小麦叶片NR活性没有明显影响。整体表现为T处理降低冬小麦叶片NR活性趋势(图3-a)。
2.2.2 大豆叶片NR活性 与CK相比,TP处理下开花期大豆叶片NR活性显著增加42.84%(P=0.023);T、P处理对大豆叶片NR活性无显著影响(图3-b)。
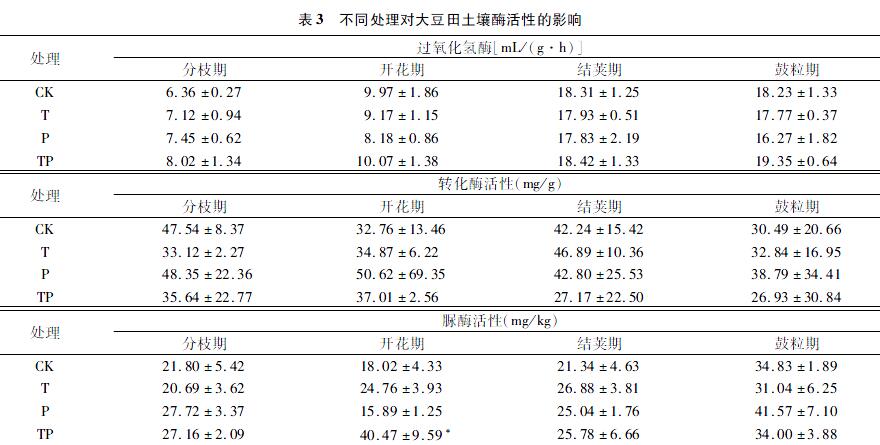
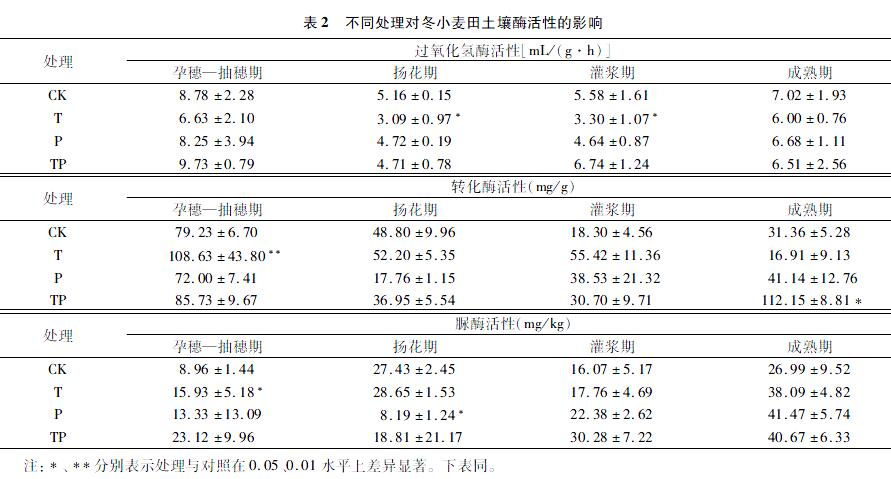
2.3 增温和降水量减少对土壤酶活性的影响
2.3.1 冬小麦土壤酶活性
2.3.1.1 过氧化氢酶活性 表2为不同处理对冬小麦土壤酶活性的影响。与CK相比,T处理下过氧化氢酶活性在扬花期显著降低40.12%(P=0.014),在灌浆期显著降低 40.86%(P=0.035);在全生育期中,P处理降低土壤过氧化氢酶活性,TP处理呈增加土壤过氧化氢酶活性,但未达到显著水平。
2.3.1.2 转化酶活性 与CK相比,T处理下孕穗—抽穗期转化酶活性极显著增加,增幅达37.11%(P=0.003);TP处理下成熟期转化酶活性极显著增加,增幅达2.58倍(P=0.009);P处理在灌浆期前降低土壤转化酶酶活性,在灌浆后增加土壤转化酶酶活性,但未达到显著水平。
2.3.1.3 脲酶活性 与CK相比,T处理下孕穗—抽穗期土壤脲酶活性显著增加77.79%(P=0.027);P处理下,扬花期酶活性显著降低70.14%(P=0.013);整体上TP处理增加脲酶活性,但未达到显著水平。
2.3.2 大豆土壤酶活性
2.3.2.1 过氧化氢酶活性 表3为不同处理对大豆土壤酶活性的影响。与CK相比,T、P、TP处理对大豆土壤过氧化氢酶活性的影响均未达到显著水平;除分枝期外,T、P处理降低过氧化氢酶活性,TP处理在整个生育期增加过氧化氢酶活性趋势。
2.3.2.2 转化酶活性 与CK相比,T、P、TP处理对转化酶活性的影响均不显著,T处理总体上增加转化酶活性。
2.3.2.3 脲酶活性 与CK相比,TP处理下开花期脲酶活性显著增加1.25倍(P=0.033);T、P处理对脲酶活性无显著影响。
3 讨论
3.1 生物量
有研究表明,增温会增强植物的光合作用和水分利用率,进而加快植物的生长速率,以致干物质得到有效累积,生物量增加[7],发育期提前。石福孙等研究表明,增温可能导致地下生物量降低[26],不同物种生物量对模拟增温的响应存在明显差异[6,27]。本研究结果表明,增温可增加豆麦作物地上生物量和总生物量,增温和增温与降水量减少复合处理的冬小麦生育期比CK提前5 d左右,这与大多以往研究结果[28]一致。
气候变暖对地上生物量的影响主要取决于降水量[20],降水量减少处理对冬小麦生物量影响不显著可能是由于南京降水全年较为充足,降水量减少30%产生的影响并不明显。大豆是豆类作物中对水分变化最敏感的一种,降水量减少处理对大豆地下生物量没有显著影响,但存在整个生育期降低地下生物量的趋势, 这与闫春娟等得出的大豆营养生长期水分亏缺根系生物量减少但差异不显著的结果[29]相同。
增温和降水量减少的协同作用与单因子对豆麦生物量影响上的相关关系具体表现为增温和降水量减少的协同作用对豆麦作物生物量的影响与某一存在显著影响作用的单因子处理结果一致,但这个一致性目前仅存在只有1个单因子处理影响显著的情况下,是否存在某一农作物受2种单因素处理显著影响,以及该情况下协同作用会有何变化,都需要进一步探讨。
3.2 酶活性
NR是由硝酸盐诱导形成的限速酶和调节酶,叶片内活性最高,但稳定性极差[30]。增温和降水量减少对NR活性主要产生间接影响,增温促进豆麦作物生长,使得叶片NR合成及活性提高;降水量减少无法满足豆麦作物正常生长需求,导致NR合成受阻、分解加速,造成NR含量下降,活性可能降低。本研究结果表明,增温有降低冬小麦叶片NR活性的趋势,可能由于温棚的封顶式增温幅度偏大,会导致微生物代谢活性下降[31]。另外,研究区域地处南京市,降水量丰富,因此降水量减少处理的效果并不明显。大豆开花期TP处理显著增加NR活性,T、P处理对NR活性没有显著影响,这表明作为豆类作物中对水分变化最为敏感的作物, 降水量可能是影响大豆开花期NR活性的关键因素。
土壤酶是土壤生态系统代谢的一类重要动力[32-33],温度持续升高会增加土壤酶的活性[34],也可能会降低土壤过氧化氢酶活性[35-36],降水量减少会降低土壤酶活性[37]。本研究发现,增温和降水量减少对冬小麦和大豆的土壤酶活性的影响大致相同,这与以往研究结果一致。增温和降水量减少的协同作用对冬小麦和大豆的土壤酶活性大多具有增加作用,这表明在不同品种间,也存在协同作用与单因素影响的差异性,但协同作用呈现的效果是否与单因子所占权重有关,还需进一步探讨。
4 结论
增温可增加豆麦地上、总生物量及大豆三叶期地下生物量;降水量减少对豆麦生物量无显著影响,整体呈现降低豆麦地下生物量的趋势;增温和降水量减少的协同作用可增加豆麦作物的地上和总生物量,对地下生物量无显著影响。
冬小麦叶片NR活性对3种处理响应不显著,增温和降水量减少的协同作用增加大豆开花期NR活性。对大豆田土壤,增温和降水量减少的协同作用增加了开花期土壤脲酶活性。对冬小麦土壤,降水量减少处理降低了扬花期土壤脲酶活性,并降低了土壤过氧化氢酶活性,但未达到显著水平;增温和降水量减少的协同处理可增加冬小麦成熟期土壤转化酶活性,但对土壤過氧化氢酶、脲酶活性没有显著影响。
参考文献:
[1]IPCC. Climate change 2013:the physical scientific basis.Contribution of working group I to the 5th assessment report of the intergovernmental panel on climate change[M]. Cambridge:cambridge university press,2013.
[2]张立波,景元书,娄伟平,等. 近50年华东地区雨日及降水量的变化特征[J]. 大气科学学报,2013,36(4):426-433.
[3]陈 健,王艺杰,胡正华,等. 增温和降水减少对冬小麦和大豆生长季土壤氮素的影响[J]. 生态环境学报,2017,26(7):1107-1113.
[4]Trenberth K E,Dai A G,Van Der Schrier G A,et al. Global warming and changes in drought[J]. Nature Climate Change,2014,4(1):17-22.
[5]Cabas J,Weersink A,Olale E. Crop yield response to economic,site and climatic variables[J]. Climatic Change,2010,101(3/4):599-616.
[6]Tack J,Barkley A,Nalley L L. Heterogeneous effects of warming and drought on selected wheat variety yields[J]. Climatic Change,2014,125(3/4):489-500.
[7]胡正华,周迎平,崔海羚,等. 昼夜增温对大豆田土壤N2O排放的影响[J]. 环境科学,2013,34(8):2961-2967.
[8]耿 婷,陈 金,张卫建,等. 全天增温对不同年代冬小麦品种生长发育及产量的影响[J]. 应用生态学报,2014,25(5):1357-1365.
[9]张 凯,王润元,王鹤龄,等. 温度升高和降水减少对半干旱区春小麦生长发育及产量的协同影响[J]. 中国生态农业学报(中英文),2019,27(3):413-421.
[10]闫艳艳. 冬春季夜间增温对冬小麦植株氮代谢和籽粒蛋白质形成的影响[J]. 麦类作物学报,2018,38(2):1-9.
[11]张志良,瞿伟菁. 植物生理学实验指导[M]. 北京:高等教育出版社,2003:35-130.
[12]周礼恺,张志明. 土壤酶活性的测定方法[J]. 土壤通报,1980(5):37-38.
[13]周广胜. 气候变化对中国农业生产影响研究展望[J]. 气象与环境科学,2015,38(1):80-94.
[14]Tian Y L,Chen J,Chen C Q,et al. Warming impacts on winter wheat phenophase and grain yield under field conditions in Yangtze Delta Plain,China[J]. Field Crops Research,2012,134(3):193-199.
[15]Liu L L,Wang E L,Zhu Y,et al. Contrasting effects of warming and autonomous breeding on single-rice productivity in China[J]. Agriculture Ecosystems & Environment,2012,149(7):20-29.
[16]Das S,Bhattacharyya P,Adhya T K. Interaction effects of elevated CO2 and temperature on microbial biomass and enzyme activities in tropical rice soils[J]. Environmental Monitoring & Assessment,2011,182(1/4):555.
[17]陈书涛,桑 琳,张 旭,等. 增温及秸秆施用对冬小麦田土壤呼吸和酶活性的影响[J]. 环境科学,2016,37(2):703-709.
[18]Hoegy P,Poll C,Marhan S,et al. Impacts of temperature increase and change in precipitation pattern on crop yield and yield quality of barley[J]. Food Chemistry,2013,136(3/4):1470-1477.
[19]Waha K,Mueller C,Rolinski S. Separate and combined effects of temperature and precipitation change on maize yields in sub-Saharan Africa for mid- to late-21st Century[J]. Global and Planetary Change,2013,106(4):1-12.
[20]Zhang T,Guo R,Gao S,et al. Responses of plant community composition and biomass production to warming and nitrogen deposition in a temperate meadow ecosystem[J]. PLoS One,2015,10(4):e0123160.
[21]Gonzalez-Meler M A,Silva L B,Diasdeoliveira E,et al. Experimental air warming of a stylosanthes capitata,vogel dominated tropical pasture affects soil respiration and nitrogen dynamics[J]. Frontiers in Plant Science,2017,8:46.
[22]Alvarez G,Shahzad T,Andanson L,et al. Catalytic power of enzymes decreases with temperature:new insights for understanding soil C cycling and microbial ecology under warming[J]. Global Change Biology,2018,24(9):4238-4250.
[23]马志良,赵文强,刘 美,等. 高寒灌丛生长季土壤转化酶与脲酶活性对增温和植物去除的响应[J]. 应用生态学报,2018,29(7):2211-2216.
[24]付 微,張兴义,赵 军,等. 模拟增温对东北黑土农田作物生长季土壤呼吸的影响[J]. 生态学杂志,2017,36(3):601-608.
[25]程徐冰,吴 军,韩士杰,等. 减少降水对长白山蒙古栎叶片生理生态特性的影响[J]. 生态学杂志,2011,30(9):1908-1914.
[26]石福孙,吴 宁,罗 鹏. 川西北亚高山草甸植物群落结构及生物量对温度升高的响应[J]. 生态学报,2008,28(11):5286-5293.
[27]刘 丹,张佳华,孟凡超,等. 不同水分和增温处理对东北玉米生长和产量的影响[J]. 生态学杂志,2013,32(11):2904-2910.
[28]Wu Z T,Dijkstra P,Koch G W,et al. Responses of terrestrial ecosystems to temperature and precipitation change:a meta-analysis of experimental manipulation[J]. Global Change Biology,2011,17(2):927-942.
[29]闫春娟,王文斌,孙旭刚,等. 干旱胁迫对大豆根系发育影响初报[J]. 大豆科学,2012,31(6):924-926,931.
[30]王效科,庄亚辉,李长生. 中国农田土壤N2O排放通量分布格局研究[J]. 生态学报,2001,21(8):1225-1232.
[31]李 娜,王根绪,高永恒,等. 模拟增温对长江源区高寒草甸土壤养分状况和生物学特性的影响研究[J]. 土壤学报,2010,47(6):1214-1224.
[32]赵 婷,张 义,赵 伟,等. 退耕还林还草工程对土壤微生物和土壤酶影响的研究进展[J]. 江苏农业科学,2018,46(1):9-14.
[33]李春龙. 汞胁迫对萝卜种子萌发、幼苗根际土壤酶活性及土壤微生物的影响[J]. 江苏农业科学,2018,46(7):142-144.
[34]Allison S D,Weintraub M N,Gartner T B,et al. Evolutionary-economic principles as regulators of soil enzyme production and ecosystem function[C]// Shukla G,Varma A. Soil enzymology. Berlin Heidelberg:Springer,2011:229-243.
[35]刘 琳,朱 霞,孙 庚,等. 模拟增温与施肥对高寒草甸土壤酶活性的影响[J]. 草业科学,2011,28(8):1405-1410.
[36]钞 然,张 东,陈雅丽,等. 模拟增温增雨对典型草原土壤酶活性的影响[J]. 干旱区研究,2018,35(5):1068-1074.
[37]Xiao W,Chen X,Jing X,et al. A meta-analysis of soil extracellular enzyme activities in response to global change[J]. Soil Biology & Biochemistry,2018,123:21-32.
收稿日期:2018-11-04
基金項目:国家自然科学基金面上项目(编号:41775152、41775151);江苏省大学生实践创新计划(编号:201810300070Y)。
作者简介:孔 瑞(1994—),女,河南驻马店人,硕士研究生,主要从事农业气象与生态研究。E-mail:923875630@qq.com。
通信作者:胡正华,博士,教授,主要从事农业应对气候变化研究。E-mail:zhhu@nuist.edu.cn。
- 工业循环水处理技术改进措施
- 高压气动电磁阀方案改进及仿真分析
- 随车起重机与自卸汽车组合介绍
- 浅谈薄壁类零件高效铣削加工方式
- 浅析曲轴加工主要机床的现状及需求
- 无卡式预拉伸自动换绳法在东庞矿的应用
- 660MW发电机组氢气泄漏原因与处理
- 螺旋齿轮加工工艺研究
- 浅谈车辆应急修理
- 配电线路继电保护的改进
- 基于液压系统大臂油缸压力的抱罐车称重系统算法的研究
- 站内轨道电路预叠加ZPW—2000电码化应用
- 电动叉车的电动助力转向应用
- 煤矿机械设备维修及管理
- 车用铝合金中铝板冲压的设计及生产的关键技术
- 电站燃煤锅炉烟气污染物超低排放技术
- 浅析机车监控装置的抗干扰对策
- 工业厂房暖通空调的节能设计
- 美军单兵用防护眼镜防雾性能的现状及发展
- 青藏铁路供电技术及运营维护管理
- 特高压输电线路连接金具的高强度材料选型
- 水电站电气设备运行维护及检修
- 浅谈建筑电气节能设计
- 斜撑杆失效案例分析
- 自卸汽车后板锁紧机构剖析
- brand
- brand advertising
- brandadvertising
- brandassociation
- brand association
- brandawareness
- brand awareness
- brandconscious
- branded
- brandequity
- brand equity
- brander
- branders'
- branders
- brand extension
- brandextension
- brandies
- brand image
- brandimage
- branding
- branding iron
- branding-irons
- guildry
- guilds
- guildships
- 跛子爬山——一步三分险
- 跛子走路—— 一脚高一脚低
- 跛子走路——一脚高一脚低
- 跛曳
- 跛立
- 跛者不忘履,盲者不忘视
- 跛脚
- 跛脚上台——立场不稳
- 跛脚儿担水——一步一步来
- 跛脚毛驴——不走正道
- 跛脚的马
- 跛脚赶马——望尘莫及
- 跛脚马上阵——没有好下场
- 跛脚马碰到瞎眼骡——难兄难弟
- 跛行
- 跛行摇摆的样子
- 跛行的样子
- 跛象扯双车——总没一边儿安稳
- 跛象扯双车——总没一边安稳
- 跛足
- 跛足从事
- 跛足曳行
- 跛跨
- 跛踦
- 跛踬