张国壁 马静 左壮

摘要:谷氨酸氨基转移酶又称谷丙转氨酶,在植物氮同化中起到关键作用。本试验以小黑杨为试材构建pROKⅡ-PnAlaAT3植物表达载体,通过农杆菌介导法转化小黑杨,获得转基因植株,发现5号转基因株系叶片中PnAlaAT3基因表达量升高,但根中表达量却显著降低。对野生型和PnAlaAT3 5号转基因株系的谷氨酰胺合成酶(GS)和谷氨酰胺-α-酮戊二酸氨基转移酶(GOGAT)活性检测结果发现,在低氮条件下,PnAlaAT3转基因株系上位叶GS活LT 较野生型植株有显著增加。结果表明,PnAlaAT3基因可以提高小黑杨上位叶中GS活力。
关键词:谷丙转氨酶基因;杨树;转基因;氮素同化;谷氨酰胺合成酶活性
中图分类号: Q785 ?文献标志码: A ?文章编号:1002-1302(2020)03-0086-04
丙氨酸氨基转移酶又称谷丙转氨酶(Alanine aminotransferase,AlaAT),在磷酸吡哆醛的存在下,该酶可催化丙氨酸和α-酮戊二酸反应,生成丙酮酸和谷氨酸[1-2]。在植物中,该酶位于细胞质中,是氮素同化的关键酶[3]。第一个植物AlaAT基因是Muench等从玉米(Zea mays)中分离得到的[4],之后相继从拟南芥、大豆、小麦和苜蓿中获得了多个编码AlaAT的基因。以拟南芥为例,Igarashi等共鉴定了4个AlaAT基因,分别命名为AOAT1~AOAT4[5],而大豆[6]和苜蓿[7]也有同样的家族成员数量。从功能上来看,在拟南芥中过表达外源物种的AlaAT基因可以提高拟南芥的氮素利用效率,而过表达大麦AlaAT基因的水稻(Oryza sativa)和油菜(Brassica nupas)的产量和生物量都发生了显著的提高[8-9]。过表达大麦AlaAT基因的甘蔗也在低氮条件下表现出了提高氮素利用率的现象[10-11]。
林木AlaAT基因研究相对较晚,2017年,Xu等从杨树中共克隆得到4个成员,分别命名为PnAlaAT1、PnAlaAT2、PnAlaAT3、PnAlaAT4,其中PnAlaAT1和PnAlaAT2主要表达在叶中,并受日节律的影响,而PnAlaAT3和PnAlaAT4表达量在根中相对较多,其中PnAlaAT3在根中表达量最高,并受外源氮素的调节而显著上调[12]。但该基因在氮素同化中是否起到关键作用尚不清楚。本研究将PnAlaAT3在杨树体内进行过表达的遗传转化,并对转化株系在不同浓度氮素下的植株生长状况进行研究,从而推断该基因在杨树氮素同化中所起的作用。
1 材料与方法
1.1 试验材料
采用小黑杨作为试验材料,取材于东北林业大学试验林场。pGEM-T Easy载体和大肠杆菌(Escherichia coli) Trans1-T1感受态细胞,购自北京全式金生物技术有限公司;双元表达载体pROKⅡ和根癌农杆菌(Agrobacterium tumefaciens)菌株EHA105,由笔者实验室保存;各种限制性内切酶、T4 DNA连接酶、反转录试剂盒,均购自TAKARA公司;引物合成委托哈尔滨博士生物科技有限公司完成。其他所需试剂均为国产分析纯。
1.2 试验方法
1.2.1 载体构建 根据杨树来源的PnAlaAT3(GenBank登录号:KT768062)设计引物PnAlaAT3-F/R:5′-GCTCTAGACCATGGCTCGTGTTTCTCTTG-3′,5′-TCCCCCGGGTCACTCGCGAAACTCCTCC-3′。下划线分别为XbaⅠ和SmaⅠ的酶切位点。以重组质粒pGEM-T Easy-AlaAT3为模板,用KOD plus高保真酶扩增目的片段,扩增产生的目的基因条带为 1 446 bp。采用XbaⅠ和SmaⅠ双酶切将目的片段连接到双元表达载体pROKⅡ中,构建植物表达载体pROKⅡ-AlaAT3(图1-A)。在构建的表达载体中,AlaAT3上游启动子为组成型强启动子CaMV 35S,终止子为NOS,卡那霉素抗性基因。采用电击法将重组表达载体pROKⅡ-PnAlaAT3转入农杆菌EHA105中。
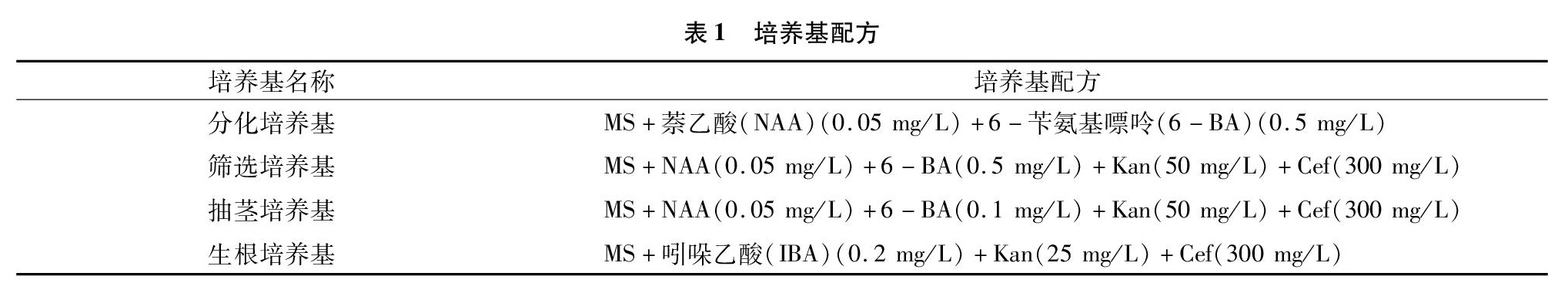
1.2.2 遗传转化 以小黑杨为受体物种,采用农杆菌介导法对其进行遗传转化。选取长势好的小黑杨无菌苗的叶片切成边长为1 cm的正方形小块,在农杆菌中侵染20 min后取出叶片,接种在不含抗生素的MS[13](Murashige & Skoog)分化培养基上,于25 ℃条件下黑暗培养2 d。之后将叶片接种到含有50 mg/L卡那霉素(Kan)和300 mg/L头孢霉素(Cef)的筛选培养基中,在25 ℃、16 h光照/8 h黑暗的条件下培养。待分化的不定芽长到1 cm左右时将其切下,转移到含有50 mg/L卡那霉素和 300 mg/L 头孢霉素的抽茎培养基中,待抗性芽长到2 cm时,将其切下并放入含有25 mg/L卡那霉素和300 mg/L头孢霉素的生根培养基中,继续培养抗性植株并生根。详细培养基配方见表1。
1.2.3 转基因植株的分子检测 以转基因小黑杨DNA为模板,用PnAlaAT3基因的特异性引物序列(PnAlaAT3-F,5′-ATGGAAGTCACTGGGTTTGG-3′;PnAlaAT3-R,5′-GAGTAGGCTGCGACAGTAAAAG-3′)进行PCR检测,目的条带送样测序。利用实时荧光定量PCR方法检测PnAlaAT3基因在转基因小黑杨中的表达情况,以PtActin基因为内参基因。PnAlaAT3定量引物序列:F:5′-GTTCCTGGCTCTGGCTTTGGG-3′;R:5′-ACTCCGTGAGACGGGAGACAACA-3′。
1.2.4 不同氮素处理及指标检测 选取健壮的转化植株和野生型无菌苗,移到灭菌的水培培养液(无蔗糖及琼脂的MS培养液)中,温室培养一段时间后,将所有试验苗分为4组,分别为野生型正常氮组(2 mmol/L NH4NO3)、转基因正常氮组(2 mmol/L NH4NO3)、野生型低氮组(1 mmol/L NH4NO3)和转基因低氮组(1 mmol/L NH4NO3)。用上述配置好的营养液装入水培装置,把小黑杨材料转入水培装置,处理2 d,处理2 d后用吸水纸吸干根部残留的营养液,收样,放入-80 ℃保存。其中叶片收样分为2个部分:第一部分是從小黑杨第1张完全展开的叶片开始依次向下取3张叶片的混样;第二部分是最下方3张叶片,分别代表小黑杨的新叶(上位叶)及老叶(下位叶)部分。转氨酶、谷氨酰胺合成酶、谷氨酸合酶等的活性用苏州科铭生物技术有限公司的试剂盒进行检测。
1.2.5 数据分析 采用SPSS 20.0软件对转基因株系与对照在表达水平及表型性状的差异显著性进行t检验(P=0.05)。所有样品为5棵苗的混合样本,生物学重复3次。
2 结果与分析
2.1 转PnAlaAT3基因的获得及分子检测
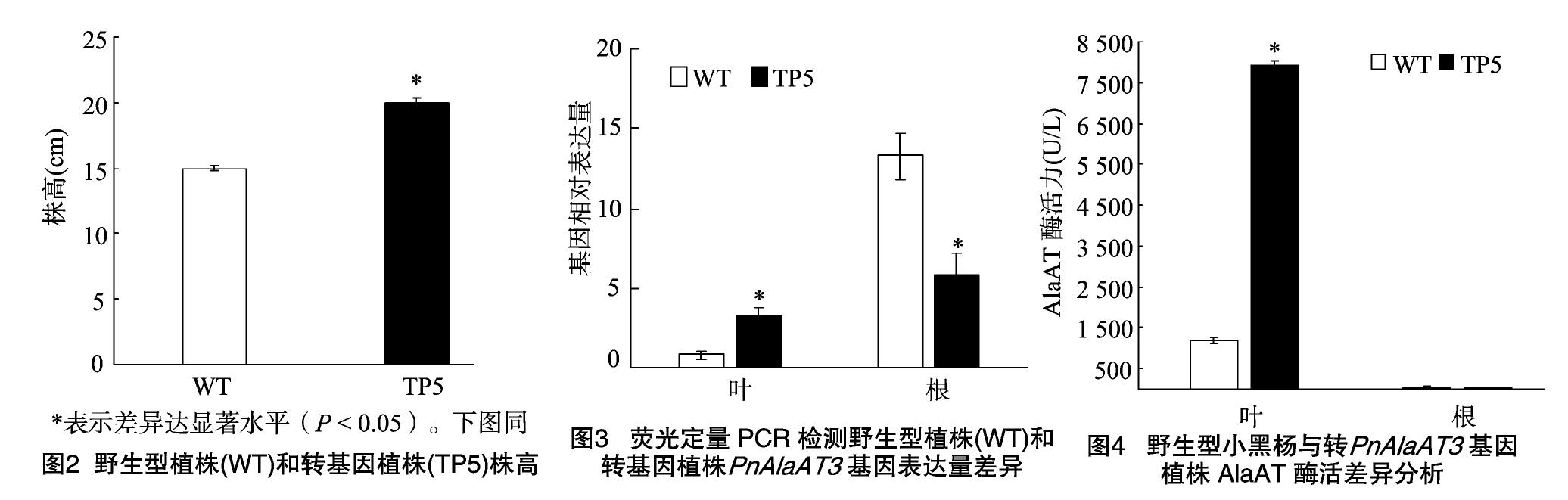
采用农杆菌介导法,将PnAlaAT3基因转入小黑杨中,共获得17棵转化株系,挑选5、6、7号转化株系(图1-B中1~3)进行DNA及RNA等后续检测,以DNA为模板的PCR检测结果表明这些条带的大小约为400 bp(图1-B),经测序确定为 354 bp,扩增结果与预期相符。这些结果初步证明PnAlaAT3基因已经成功整合到野生型小黑杨的基因组中。将5号转化株系进行无性扩繁后检测株高,结果如图 1-C和图2所示,野生型平均株高为15 cm,转化株系平均株高为20 cm,两者株高差异显著(P<0.05)。通过荧光定量PCR方法检测PnAlaAT3基因在该转化株系根和叶中的表达量,结果如图3所示,在叶片中,转基因小黑杨叶片中的基因表达量高于非转基因的植株;而在根中,转基因小黑杨中PnAlaAT3基因的表达量显著低于野生型植株。由此表明,PnAlaAT3基因不仅成功整合到野生型小黑杨的基因组中,并且在小黑杨不同组织中成功表达。
2.2 转PnAlaAT3基因植株AlaAT酶活变化
为了确定转PnAlaAT3基因的转化株系在酶活水平的变化情况,对野生型及转化株系地上和地下部分酶活进行检测,结果如图4所示,在根中,转基因植株的AlaAT酶活与野生型植株基本一致,但在叶片中转基因植株的AlaAT酶活相对于野生型植株有显著的提高。
2.3 转PnAlaAT3基因小黑杨对氮同化相关基因酶活的影响
GS和GOGAT是氮素同化过程中关键的基因,本试验对不同氮素下转PnAlaAT3基因株系的谷氨酰胺合成酶(GS)以及谷氨酰受-α-酮戊二酸氨基轉移酶(GOGAT)活力进行检测,结果如图5所示,在根中以及下位叶中,GS以及GOGAT活力没有显著差异,即使在不同氮素处理的条件下也是如此;在上位叶中,转PnAlaAT3基因的转化株系GOGAT活力没有显著区别,但GS活力在低氮条件下相对于野生型植株有明显的提高,且差异达到显著水平。
3 结论与讨论
在植物中,谷丙转氨酶不仅在逆境条件下发挥一定的作用,在外界氮素发生变化时,该酶也同时起到提高氮素利用的功能[13]。有研究表明,在小麦中氮素利用率的高低与谷丙转氨酶表达量成正相关[14-15]。在油菜中组成型表达大麦的谷丙转氨酶基因,结果导致在低氮条件下油菜的氮素利用率提高[9,16]。将大麦AlaAT基因转入水稻,并加以水稻的根部特异性启动子,结果导致转基因水稻分蘖数明显增加,根更加稠密[8]。同时,转基因水稻地上
部分的生物量和含氮总量也显著增加,籽粒产量显著高于野生型水稻[8]。同时,谷氨酸、谷氨酰胺和天冬氨酸等的含量也发生相应变化[17]。
本研究中将PnAlaAT3基因转入野生型小黑杨,获得了过表达株系,在相同生长条件下发现转基因植株长势优于野生型植株,株高有显著增加。且转基因植株叶片中的AlaAT酶活高于野生型植株叶片中的AlaAT酶活。在低氮处理下,转化株系的上位叶片中GS活力显著高于野生型植株,株高也优于野生型植株。GS活力升高会积累更多的谷氨酰胺,以提供给上位叶更多的原材料,从而使转基因植株长势更好。
参考文献:
[1]Miyashita Y,Dolferus R,Ismond K P,et al. Alanine aminotransferase catalyses the breakdown of alanine after hypoxia in Arabidopsis thaliana[J]. The Plant Journal,2007,49(6):1108-1121.
[2]Yonaha K,Suzuki K,Toyama S. Streptomyces beta-alanine:alpha-ketoglutarate aminotransferase,a novel omega-amino acid transaminase. Purification,crystallization,and enzymologic properties[J]. Journal of Biological Chemistry,1985,260:3265-3268.
[3]黄冰艳,高 伟,苗利娟,等. 谷氨酰胺合成酶基因研究进展及其在植物氮代谢调控中的应用[J]. 中国农学通报,2010,26(23):53-57.
[4]Muench D G,Christopher M E,Good A G. Cloning and expression of a hypoxic and nitrogen inducible maize alanine aminotransferase gene[J]. Physiologia Plantarum,1998,103(4):503-512.
[5]Igarashi D,Miwa T,Seki M,et al. Identification of photorespiratory glutamate:glyoxylate aminotransferase (GGAT) gene in Arabidopsis[J]. The Plant Journal,2003,33(6):975-987.
[6]Rocha M,Sodek L,Licausi F,et al. Analysis of alanine aminotransferase in various organs of soybean (Glycine max) and in dependence of different nitrogen fertilisers during hypoxic stress[J]. Amino Acids,2010,39(4):1043-1053.
[7]Ricoult C,Echeverria L O,Cliquet J B,et al. Characterization of alanine aminotransferase (AlaAT) multigene family and hypoxic response in young seedlings of the model legume Medicago truncatula[J]. Journal of Experimental Botany,2006,57(12):3079-3089.
[8]Shrawat A K,Carroll R T,DePauw M,et al. Genetic engineering of improved nitrogen use efficiency in rice by the tissue-specific expression of alanine aminotransferase[J]. Plant Biotechnology Journal,2008,6(7):722-732.
[9]Good A G,Johnson S J,De Pauw M,et al. Engineering nitrogen use efficiency with alanine aminotransferase[J]. Revue Canadienne De Botanique,2007,85:252-262.
[10]Snyman S J,Hajari E,Watt M P,et al. Improved nitrogen use efficiency in transgenic sugarcane:phenotypic assessment in a pot trial under low nitrogen conditions[J]. Plant Cell Reports,2015,34(5):667-669.
[11]徐红卫,郭桂梅,黄赛华, 等. 两份大麦纯合突变体氮代谢相关基因于不同生长发育时期的表达[J]. 植物生理学报,2017,53(11):2031-2036.
[12]Xu Z,Jing M,Qu C,et al. Identification and expression analyses of the alanine aminotransferase(AlaAT) gene family in poplar seedlings[J]. Scientific Reports,2017,7:45933.
[13]Enosawa S,Dozen M,Tada Y,et al. Electron therapy attenuated elevated alanine aminotransferase and oxidative stress values in type 2 diabetes-induced nonalcoholic steatohepatitis of rats[J]. Cell Medicine,2013,6(1/2):63-73.
[14]Tian H,Fu J,Drijber RA,et al. Expression patterns of five genes involved in nitrogen metabolism in two winter wheat(Triticum aestivum L.) genotypes with high and low nitrogen utilization efficiencies[J]. Journal of Cereal Science,2015,61:48-54.
[15]董召娣,易 媛,張明伟,等. 春性和半冬性小麦花后旗叶和籽粒氮代谢关键酶活性的差异[J]. 麦类作物学报,2015,35(8):1098-1106.
[16]Mauchline T H,Fowler J E,East A K,et al. Mapping the Sinorhizobium meliloti 1021 solute-binding protein-dependent transportome[J]. Proceedings of the National Academy of Sciences of the United States of America,2006,103(47):17933-17938.
[17]Kan C C,Chung T Y,Juo Y A,et al. Glutamine rapidly induces the expression of key transcription factor genes involved in nitrogen and stress responses in rice roots[J]. BMC Genomics,2015,16(1):731.
- 让语文作业设计个性飞扬
- 让拓展趣味化,让学习深度化
- 提升小组讨论效率“四策略”
- 图文结合,优化语文教学
- 谈核心素养下文本研读的着眼点
- 例说课堂教学中的当场检测
- 让学生在童话中尽情体验
- 浅谈提高学生阅读速度的方法
- 课堂因生成而美丽
- 寻求随文练笔落脚点,助力语用能力提升
- 让学生在阅读期待中走向“和美”
- 以对话推进个性朗读
- 巧辟蹊径,在创意读写中发展核心能力
- 朗读
- 让学引思,阅读教学的有效策略
- 说理文教学方法探究
- 个性化阅读教学策略探微
- 基于儿童的言语实践活动浅探
- 立足学生视角,构建灵动语文课堂
- 儿童阅读能力发展有效策略浅探
- 提炼运用,让文本信息资源价值最大化
- 根据语文要素,展开语用训练
- 根据农村实际,提升课外阅读整体效益
- 聚焦延展,为学生言语智能发展助力
- 运用单元整合策略,让语文课堂更出彩
- rescuing
- resculpt
- resculpted
- resculpting
- resculpts
- resculpture
- resculptured
- resculptures
- resculpturing
- resealable
- resealed
- resealing
- reseals
- research
- researchable
- researchanddevelopment
- research and development
- researched
- researcher
- researcher, researchist
- researcher's
- researchers
- researches
- researchful
- researching
- 忠顺
- 忠顺亲王
- 忠驱义感
- 忠骨
- 忠魂
- 忠魂毅魄
- 忠鲠
- 忡
- 忡忡
- 忡忡忧心
- 忡怅
- 忡怛
- 忡惕
- 忡然
- 忣
- 忣忣
- 忤
- 忤嫚
- 忤往
- 忤怨
- 忤恨
- 忤意
- 忤慢
- 忤时
- 忤物