梁芳 刘冰 檀小辉 赵仕花 梁泽锐 苏丽星 陈飞凤 梁梦雅
摘要:以三年生红花玉蕊为研究对象,通过JN-G02型高智能多参数土壤肥料养分检测仪测定其新梢生长发育过程中不同部位矿质元素(氮、磷、钾、铁、铜)的含量,明确各元素在植株不同部位的分布规律,探索此过程中影响植株各部位发育的关键矿质元素。结果表明,红花玉蕊新梢发育过程中成熟叶及老叶中的N、P含量在新梢生长前期呈迅速下降趋势(老叶P元素除外),茎尖及幼嫩茎段的N含量在新梢生长前期也呈下降趋势,但半木质化茎段的N含量呈上升趋势。叶片中的K、Fe、Cu含量在新梢生长前期总体呈现上升趋势,幼嫩茎段和半木质化茎段的K含量在生长后期达到最大值。叶片、茎段的N含量与P、K、Fe、Cu含量均呈负相关,其中N含量与P含量均呈极显著负相关(P<0.01),P、K、Fe、Cu含量之间呈正相关,其中叶片的P含量与Fe含量呈显著正相关(P<0.05)。可见,氮、磷、钾、铁、铜矿质元素对红花玉蕊新梢的发育有着重要影响,在养护管理施肥方面应该注意在不同时间根据不同部位补充各矿质元素,尤其应关注植株新稍发育期氮含量与磷出现极显著负相关现象,注意氮、磷元素用量的协调,同时要合理搭配其他必要的矿质元素,以保证红花玉蕊植株营养均衡、健康、生长迅速,使红花玉蕊施肥达到合理化和规范化。
关键词:红花玉蕊;新梢;矿质元素;时空变化
中图分类号:S685.01?文献标志码:A?文章编号:1002-1302(2020)21-0133-07
红花玉蕊(Barringtonia acutangula)是玉蕊科红花玉蕊属的常绿小乔木植物,原产于东南亚海滨地带。Abhik研究发现,红花玉蕊具有一定的耐旱和抗涝能力,是印度阿萨姆邦南部巴拉克谷洪泛平原植被中的优势树种,能形成天然林以长时間抵御洪水淹浸,也是该区常见的沼泽地树种[1]。该树种株形美观,叶大常绿,花序下垂,花色深红,极具园林观赏价值,已在我国成功引种[2]。矿质元素是植物必需营养物质,是树体生长发育不可或缺的物质基础。目前,大多数植物种植仍采用经验法施肥,存在盲目性和不确定性,与精准林业的发展要求不适应。对于红江橙(Hongjiang orange)[3]、梨枣(pear-jujube)[4]、越橘(Vaccinium vitis-idaea)[5]、油茶(Camellia oleifera)[6]和甜柿(Diospyros kaki)[7]等植物矿质营养动态变化的研究,研究对象主要为发育过程中的叶片,而植物新梢的生长状况直接影响树体生长、发育和开花结果。笔者在前期课题研究中已初步揭示了红花玉蕊在淹水、盐害等逆境胁迫下的生理生化响应机制,但仍有较多问题尚未明确,使其难以进行大规模人工驯化栽培,关于新梢发育过程中矿质元素变化特性的研究目前尚未见报道[8-10]。本研究通过对红花玉蕊新梢发育过程中矿质元素含量进行时空变化分析,明确红花玉蕊新梢发育过程中不同部位各元素含量的变化规律,找出新梢发育过程中影响各个部位不同时期生长发育的关键矿质元素,明确各元素之间的相互关系,这对深入了解红花玉蕊树体对营养元素的需求动态、吸收能力和养分调控等具有重要现实意义,同时也可为红花玉蕊植株的合理施肥、科学养护提供理论依据。
1 材料与方法
1.1 材料选取
以玉林师范学院温室大棚内三年生红花玉蕊实生苗为试材,选取生长正常,质量与大小基本一致,株高约为40 cm且无病虫害的红花玉蕊幼苗用于试验。采样时间为2019年4月10日至2019年6月18日。采样期间温室大棚内最高温度为 46.7 ℃,最低温度为19.3 ℃,平均温度为33 ℃,最高湿度为99.9%,最低湿度为39.2%,平均湿度为69.55%。
1.2 试验方法
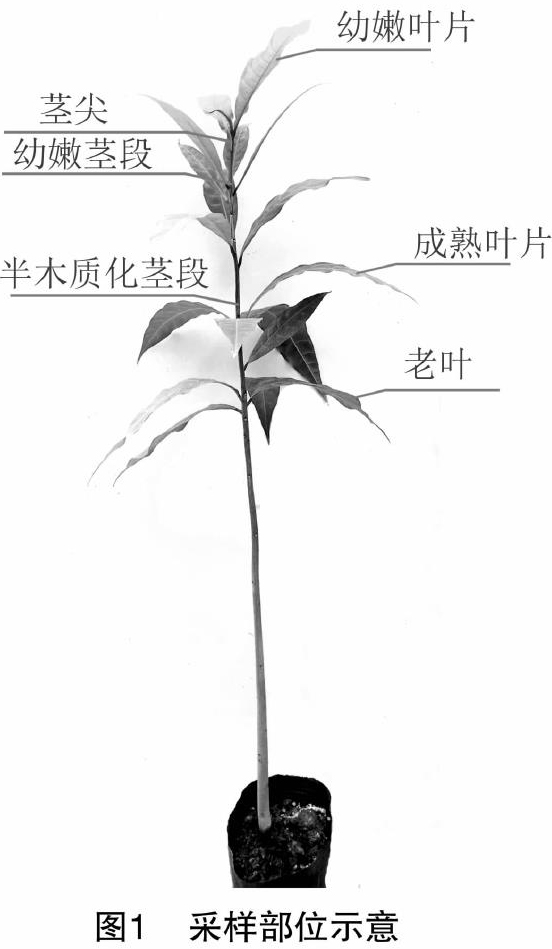
1.2.1 样本调查方法 每2个星期选取15~25株生长健壮的红花玉蕊新梢茎尖(新梢茎尖生长点向下3 cm以内)、幼嫩茎段(茎尖下组织幼嫩、未木质化的茎段)、半木质化茎段(幼嫩茎段下柔韧性好的茎段)、幼嫩叶片、成熟叶片、老叶共6个部位(图1)进行取样,各部位分别混合为1份样品,3次重复,应在植株外围不同方向(东西南北)进行样品采集,测定植株不同部位的矿质元素N、P、K、Fe、Cu含量。用干布擦净样品。并将其剪成1~2 mm碎片,混匀后放入研钵中研磨至有汁液浸出,然后用低速离心机离心,用胶头滴管吸取上清汁液2滴置于10 mL量筒中,加水至10 mL,即为待测液(100倍稀释汁液)混匀备用。根据要测定元素的不同分别加入不同药品进行反应,待反应结束后立即使用高智能多参数土壤肥料养分检测仪(郑州锦农科技有限公司,锦农牌JN-G02型)测定N、P、K元素含量。用胶头滴管取上清汁液4滴置于10 mL量筒中,加水至10 mL,即为待测液,混匀备用。根据要测定元素的不同分别加入不同药品试剂进行反应,反应结束后即可上机测定Fe、Cu元素含量。
1.2.2 数据处理 采用Excel 2007进行试验数据初步整理,用GraphPad Prism 7.00作图,用SPSS 23.0软件对茎尖、幼嫩茎段、半木质化茎段、幼嫩叶片、成熟叶片和老叶的各矿质元素含量进行单因素方差分析及相关性分析,采用邓肯氏多重比较法进行差异显著性分析。
2 结果与分析
2.1 硝态氮含量的变化
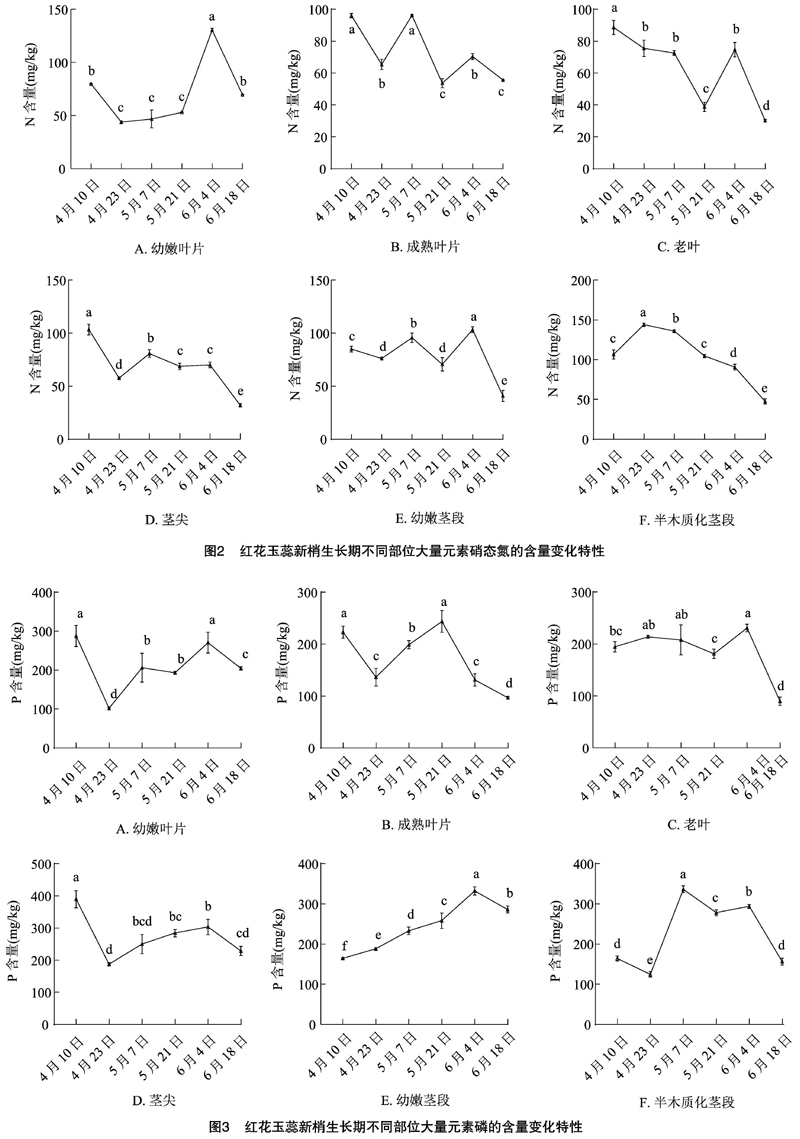
从图2可以看出,红花玉蕊植株成熟叶片及老叶中N含量整体呈下降趋势,在4月10日最大,其中老叶N含量与其他采样时间呈显著差异(P<0.05);幼嫩叶片的N含量在6月4日达到峰值,与其他时间均呈显著差异(P<0.05),且高于成熟叶片和老叶;茎尖N含量在新梢发育初期(4月10日)与其他时间呈显著差异(P<0.05),半木质化茎段的N含量随新梢的生长发育呈先上升后下降的趋势;6月18日茎段各部位的N含量均降至最低值,此时各茎段部位N含量与其他时间均呈显著差异(P<0.05)。可见,不同发育时期各个部位的N含量差异较大。
2.2 磷元素含量的变化
新梢发育过程中红花玉蕊植株6个部位的P元素含量随时间的变化有升有降(图3),新梢发育初期(4月10日)幼嫩叶片的P含量在叶片中最高,且与其他时间(除6月4日外)呈显著差异(P<0.05)。新梢发育后期(6月18日)幼嫩叶片P含量较成熟叶片、老叶高,此时成熟叶片、老叶的P含量显著低于其他时间(P<0.05)。在新梢发育初期(4月10日)茎尖的P含量显著高于中后期(P<0.05)。幼嫩茎段P含量随生长时期的推移呈先上升后下降趋势,在6月4日达到峰值,与其他时间呈显著差异(P<0.05)。在5月7日半木质化茎段的P含量达到峰值,与其他时间呈显著差异(P<0.05)。
2.3 钾元素含量的变化
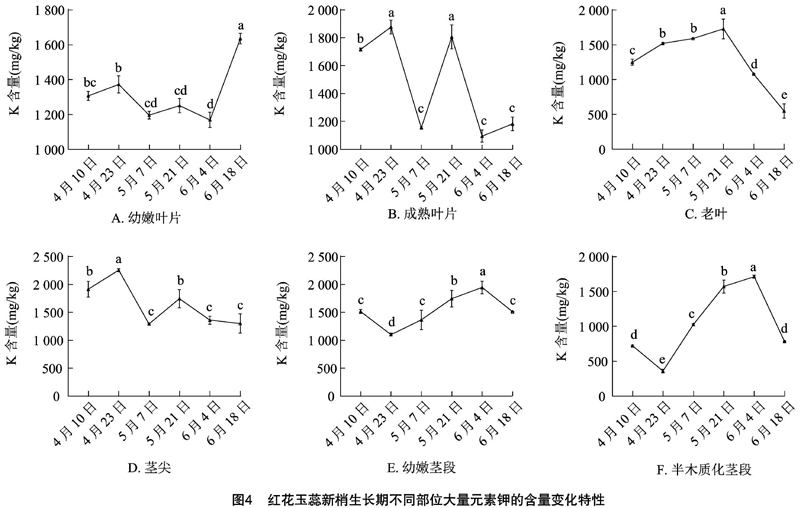
新梢发育过程中红花玉蕊植株6个部位的K元素含量整体变化不一(图4),幼嫩叶片的K含量在生长后期(6月18日)达到最大值,与生长前中期呈显著差异(P<0.05),且均高于同时期成熟叶片和老叶。老叶的K元素含量整体呈明显的先升后降趋势,生长前中期含量显著高于生长后期(P<0.05)。4月23日茎尖的K含量显著高于生长中后期(P<0.05)。幼嫩茎段和半木质化茎段的K含量均在6月4日达到峰值,且显著高于前期(P<0.05)。可见K元素含量在老叶、幼嫩茎段和半木质化茎段中的变化趋势更明显。
2.4 铁元素含量变化
新梢发育过程中红花玉蕊植株Fe含量除幼嫩茎段整体呈下降趋势外,其他部位在生长前期均有所升高(图5)。幼嫩叶片的Fe含量在5月7日达到峰值,与其他时间均呈显著差异;成熟叶片和老叶的Fe含量峰值出现在4月23日。茎尖Fe含量在生长中期较高,半木质化茎段中的Fe元素含量峰值出现在新梢生长前中期(4月23日至5月7日),显著高于其他时间。可见Fe元素含量与叶片及茎段的老化程度密切相关,同时在生长后期均呈较低水平。
2.5 铜元素含量变化
红花玉蕊植株6个部位的Cu元素含量总体上在新梢发育前期呈逐渐上升趋势(图6)。幼嫩叶片和成熟叶片的Cu含量在各生长时期变化较相似,在前中期含量较高,最低值均出现在6月4日,但老叶的Cu含量在6月4日却达到了峰值,与其他时间呈显著差异(P<0.05)。可见红花玉蕊叶片的Cu含量随着叶片成熟程度变化较大。幼嫩茎段及半木质化茎段的Cu含量在4月23日达到最大值,显著高于其他时间(P<0.05),且幼嫩茎段和半木质化茎段在新稍生长中后期Cu含量整体均呈下降趋势,但茎尖Cu含量在5月7日达到最大值,与其他时间呈显著差异(P<0.05)。可见,茎中Cu含量随新稍发育时期变化较大。
2.6 新梢生长发育过程中茎叶部位矿质元素含量之间的相互关系
在红花玉蕊新梢生长发育过程中,新梢的叶片和茎段矿质元素含量的相关性分析结果见表1。叶片的N含量与P、K、Fe、Cu含量呈负相关,其中与P含量呈极显著负相关(P<0.01),N含量与Fe含量呈显著负相关(P<0.05),P含量与K、Fe、Cu含量呈正相关,其中与Fe含量呈显著正相关(P<0.05)。K、Fe、Cu元素含量之间呈正相关。茎段的N含量与P、K、Fe、Cu含量呈负相关,其中与P含量呈极显著负相关(P<0.01),P、K、Fe、Cu元素含量之间呈正相关。
3 结论与讨论
大量元素N是植株蛋白质、磷脂、酶、叶绿素、生物膜等的重要组成成分。P元素是植物激素、核酸、磷脂、酶等的重要组成成分,可促进糖运转。K元素是植物体内酶的活化剂,可促进光合作用、糖代谢、脂肪代谢及蛋白质合成,提高植物的抗寒性和抗倒伏能力,一般以离子状态存在于植物体内,调节K离子浓度可提高细胞渗透压,从而有利于植物对水分的吸收,进而提高植株的抗旱能力[11]。微量元素Fe是叶绿素合成所必需的矿质元素,参与植物体内氧化还原反应、电子传递及呼吸作用[12]。微量元素Cu是植物体内多种氧化酶的重要组成成分,在氧化还原反应中起重要作用,同时影响植株对Fe的利用及固氮作用。另外,Cu与叶绿素的形成有直接關系,具有提高叶绿素稳定性的作用[13-14]。
试验结果表明,红花玉蕊新梢发育过程中成熟叶片及老叶N含量在生长前期最大,成熟叶片、老叶及茎段各部位N含量均在生长后期达到了最低值,在生长后期幼嫩叶片的N含量显著大于成熟叶和老叶;新梢发育前期幼嫩叶片及茎尖中的P含量最高,但幼嫩叶片的K含量在生长后期出现最大值。可见,新梢生长发育初期红花玉蕊树体需要消耗大量N与P,因为此时期新梢和树高的加速生长均需要消耗大量的矿质营养元素。到生长中后期,植株新梢生长发育速度减缓,营养器官发达,植株地上部分吸收养分能力强,吸收的养分可以供红花玉蕊生长,因此,该时期红花玉蕊植株对N和P的需求量减少。叶片中大量元素K和微量元素Fe、Cu的含量在新梢发育初期总体上呈迅速上升趋势,中后期明显下降,说明新梢生长前期红花玉蕊树体对K和Fe、Cu元素的需求量也较大,以提高植物叶片光合能力。
文冠果(Xanthocerassorbifolia)在新梢生长发育初期需要消耗大量元素N、P、K与微量元素Fe,在新梢生长初期茎段中Cu含量低于叶片,生长后期茎段中Cu含量高于叶片;在新梢生长初期茎段中K元素含量较叶片高,而生长后期茎段K元素含量则低于叶片[15]。酥梨(Suli pear)叶片N、P、K、Cu的含量随叶龄的增加而降低[16]。核桃(Juglans regia)叶片中N、P、K含量在整个生育期内总体上呈逐渐降低趋势[17]。香榧(Torreya grandis)在果实膨大初期,叶片中N、P、K含量均有不同程度的下降,而Cu元素含量在整个试验过程中均呈现上升趋势[18]。黄连木(Pistacia chinensis)随季节性变化,叶片的N、P、K含量总体呈下降趋势[19]。四季柚(Citrus maxima)生长期叶片N、P、Fe元素含量逐渐下降,K元素含量表现先增后降趋势,Cu元素含量则总体表现稳定[20]。综上所述,红花玉蕊新梢生长发育过程中成熟叶及老叶N、P含量下降,其变化趋势与大多数植物相似,Fe、Cu含量随着新梢发育进程总体上呈上升趋势,幼嫩茎段除外。
红花玉蕊矿质元素含量变化的相关性分析结果表明,在新梢发育过程中叶片、茎段的N含量与P、K、Fe、Cu含量均呈負相关,其中N含量与P含量均呈极显著负相关,说明它们之间存在拮抗作用,叶片、茎段中的N元素显著抑制P元素的吸收。在日本栗(Castanea crenata)叶片矿质元素含量的变化中,N含量与K含量呈显著正相关[21];枣树(Ziziphus jujube)叶片中K含量与Fe、Cu含量呈正相关[22];黄金梨(Pyrus pyrifoliaNakai cv. ‘Whangkeumbae)的N含量和P含量在叶片和果实中均呈显著正相关[23];关于红枣(Ziziphus zizyphus)叶片中矿质元素含量动态变化的研究表明,在整个生育期中叶片N含量和P含量呈下降趋势,K含量呈先升后降趋势,其中N含量和P含量间呈极显著正相关[24];木瓜杏(Armeniaca vulgaris)叶片Fe含量和Cu含量呈极显著正相关[25];库尔勒香梨(Pyrus sinkiangensis)叶片Fe含量和Cu含量之间呈现显著正相关[26]。综上所述,红花玉蕊与大多数植物K、Fe、Cu元素含量之间的相关性相似,但N含量和P含量之间的相关性与大多数植物的研究结果相反,呈负相关关系。
综上分析可见,N、P、K、Fe、Cu等矿质元素对红花玉蕊新梢的发育有着重要影响,在养护管理施肥方面应该注意在不同时间根据不同部位补充这些矿质元素,尤其应关注植株新稍发育期N含量与P含量出现极显著负相关的现象,注意N、P元素用量的协调,同时要合理搭配其他必要的矿质元素,保证红花玉蕊植株营养均衡,才能保证其健康、迅速生长,使红花玉蕊施肥达到合理化和规范化。
参考文献:
[1]Abhik G. Water availability,poverty and socio-economic crisis in the floodplains of Barak Valley,Assam,North East India[D]. Assam:Assam University,2006.
[2]宋莉英,黎昌汉. 珍奇的庭园树木——4种外来红花玉蕊科植物[J]. 广东园林,2009,31(3):51-53.
[3]杨少瑕,朱银玲. 红江橙叶片矿质元素含量的测定及元素分布状态分析[J]. 汕头大学学报(自然科学版),2019,34(2):47-51.
[4]黄丽萍,帅 芳,张 正,等. 梨枣叶片内矿质元素年动态变化研究[J]. 山西农业科学,2017,45(2):194-196,226.
[5]周 强,耿佳麒,王宇航,等. 越橘花芽分化期叶片矿质元素含量和C/N变化分析[J]. 东北农业大学学报,2017,48(2):37-45.
[6]严江勤,曹永庆,姚小华,等. 油茶春梢发育期叶片和果实中氮磷钾元素的动态变化[J]. 中南林业科技大学学报,2016,36(2):50-55.
[7]宋少华,刘 勤,蒋振莹,等. 甜柿叶片矿质元素含量的年周期变化规律[J]. 扬州大学学报(农业与生命科学版),2016,37(1):108-112.
[8]梁 芳,黄寿镕,於艳萍,等. 红花玉蕊对淡水全淹胁迫的生长及生理响应[J]. 西南林业大学学报,2019,39(3):18-25.
[9]梁 芳,卜小英,黄秋伟,等. 水淹对红花玉蕊幼苗生长及生理特性的影响[J]. 西北林学院学报,2019,34(2):48-55.
[10]梁 芳,黄秋伟,檀小辉,等. 潮汐系统下不同盐度水体对红花玉蕊幼苗生理特性的影响[J]. 南方农业学报,2018,49(11):2250-2255.
[11]姚玉霞,李泽鸿,曹 杰,等. 几种主要营养成分氮、磷、钾在烟草生长中的作用[J]. 农业与技术,1995(3):43-45.
[12]白 洁. 微量元素在作物生长中的作用[J]. 农村科技,2007(9):22.
[13]王慧芳,张凤杰,那日苏,等. 铜胁迫对玉米幼苗生长的影响[J]. 安徽农业科学,2016,44(9):60-61.
[14]朱 健,范菲菲,赵丽华,等. 铜对海菜花种子萌发及幼苗生长的影响[J]. 西部林业科学,2016,45(2):135-139.
[15]梁 芳,卫旭芳,白永超,等. 文冠果新梢发育过程中不同部位矿质元素的变化特性[J]. 浙江农林大学学报,2018,35(4):624-634.
[16]闫 敏,李 磊,霍晓兰,等. 酥梨营养器官中矿质元素周年动态变化[J]. 山西农业科学,2011,39(8):800-802.
[17]宫永红,盖素芬,赵宝军,等. 核桃叶片中氮、磷、钾含量及年动态变化规律研究[J]. 辽宁林业科技,2002(6):9-10,21.
[18]刘萌萌,曾燕如,江建斌,等. 香榧叶片中8种矿质元素年周期季节性变化规律[J]. 经济林研究,2014,32(2):105-109.
[19]王 丹. 黄连木叶片矿质元素季节性变化规律研究[D]. 重庆:西南大学,2012.
[20]陈 巍,郭秀珠,黄品湖,等. 四季柚生长期矿质元素含量的动态变化[J]. 园艺学报,2011,38(增刊1):64.
[21]郑瑞杰,王德永,雷 鸣. 日本栗叶片矿质营养元素含量年动态变化的研究[J]. 西北林学院学报,2008,23(4):14-17.
[22]宋艳波,杨佩芳,李六林,等. 枣树年周期中叶片矿质元素含量动态变化及其相关性研究[J]. 山西农业大学学报(自然科学版),2006,26(4):340-341,390.
[23]林敏娟,王振磊,徐继忠,等. “黄金梨”叶片及果实中主要矿质元素量变化及相关关系[J]. 北方园艺,2011(19):5-7.
[24]黄 岳,岳海英. 日光温室长红枣叶片矿质元素动态变化[J]. 农业与技术,2015,35(22):47.
[25]景 淼,翟明普,高香玲,等. 木瓜杏叶片中矿质元素含量的动态变化研究[J]. 中国农学通报,2011,27(8):199-203.
[26]朱海峰,马建江,刘 艳,等. 不同产量库尔勒香梨叶片中微量矿质元素含量的年变化研究[J]. 新疆农业科学,2017,54(5):843-852.王宏辉,顾俊杰,房伟民,等. 2种不同耐热性的盆栽红掌品种抗性机制比较[J]. 江苏农业科学,2020,48(21):140-145.
- 新形势下理工科高校人文社科教育探究
- 中国传统生态智慧在高等教育可持续发展中的现代价值
- 江西高校校园电视台创新发展研究
- 深度产教融合背景下应用型人才培养探索
- 新形势下高校安全保卫联勤联动机制研究
- 笛卡尔哲学思想和方法论启迪下的大学生创新创业项目培育与优化
- 高校贫困生资助精准化研究
- 应然与实然:高校辅导员职业的两个维度
- 应用技术型高校师资队伍建设
- 翻转课堂在“ERP软件”课程中的应用初探
- “问题链导学”教学模式在“中国近现代史纲要”课的应用
- 社会科学方法论对高校思政课教学的启示
- 医学院校(5+3)研究生实验课教学几点思考
- 大数据时代基于在线自动评改的英语写作反馈研究
- 山西应用型高校思政实践教学研究
- 新媒体时代茶文化融入英语教学研究
- “思想道德修养与法律基础”课的“一微一台”式教学
- PBL教学法在返校实习生教学中的实践探索
- 基于核心素养的高等数学教学改革探究
- 双语教学考核体系改革与实践
- 应用型民办高校思政课教法转变为学法的教学改革研究
- “质量管理与控制”课程教学改革与思考
- 课程思政理念下概率论与数理统计教学改革的探索与实践
- 汉语言文学专业写作类课程教学改革探讨
- 专业剖析、说课的主要问题与改进策略
- unshrillest
- unshrined
- unshrinkingly
- unshrived
- unshriveled
- unshrivelled
- unshriven
- unshrubbed
- unshrugging
- unshrunk
- unshrunken
- unshuffled
- unshunnable
- unshunted
- unshut
- unshuttered
- unshy
- unsickened
- unsiding
- unsidling
- unsieged
- unsieved
- unsighing
- unsightlier
- unsightliest
- 焱攸
- 焱炎
- 焱烘烘
- 焱焰
- 焱焱
- 焱起
- 焱锐猱狞
- 焱集
- 焱飞
- 焱鸷
- 然
- 然且
- 然乃
- 然凭
- 然凭地
- 然则
- 然即
- 然却
- 然可
- 然后
- 然否
- 然否问
- 然如
- 然後
- 然或