王苗苗 梅肖乐 冯子偃 徐建荣 韩晓磊


摘要:通过多元统计分析和线粒体DNA测序技术,对中华鳖地方特色品种——太湖花鳖雌雄群体的形态和遗传差异进行了分析研究。结果显示,在形态上,对太湖花鳖雌雄群体各项形态参数进行t检验差异分析,发现雄性尾部的生长与其体质量的增加相关显著,呈线性关系;对太湖花鳖雌雄群体形态参数进行主成分分析,并作出主成分1和主成分2的散点图,得出雌雄群体形态差异不明显。对太湖花鳖雌雄群体线粒体D-loop控制区序列进行差异分析,雌雄群体间遗传差异为0.0114;构建N-J聚类系统树,雌雄群体无单独聚类,群体遗传差异不明显。结果表明,太湖花鳖个体形态差异与其性别存在一定程度的相关性,特别是尾部特征,可用于雌雄性别的初步判断,但雌雄群体在外观形态和线粒体DNA上均没有明显差异。
关键词:太湖花鳖;多元统计分析;D-loop控制区;形态差异;遗传差异
中图分类号:S966.5文献标志码:A
文章编号:1002-1302(2021)01-0142-04
作者简介:王苗苗(1987—),女,江苏南京人,硕士,水产工程师,主要从事水生生物学研究和水产技术推广工作。E-mail:miaomiaotgz@163.com。
通信作者:韩晓磊,硕士,高级实验师,从事淡水水生生物学研究。E-mail:84125241@qq.com。
多元统计分析可根据形态特征的多个参数,对多个互相关联的指标和对象进行统计分析以揭示其规律,符合农业科学的研究特征[1]。目前,多元统计分析已在水生动物的物种种质评定、种群遗传变异、性别差异分析等领域开展了大量的研究,然而对龟鳖类物种的研究还鲜有报道[2-6]。线粒体DNA作为细胞中遗传信息的重要载体,具有长度短、含量高、进化速度快和母性遗传等特点,特别是D-loop控制区,因其不编码蛋白,承受选择压力较小,是线粒体DNA中进化最快的部分,已成为水生动物种类鉴定、遗传多样性、遗传变异、系统进化、性别差异等方面的研究热点[7-13]。
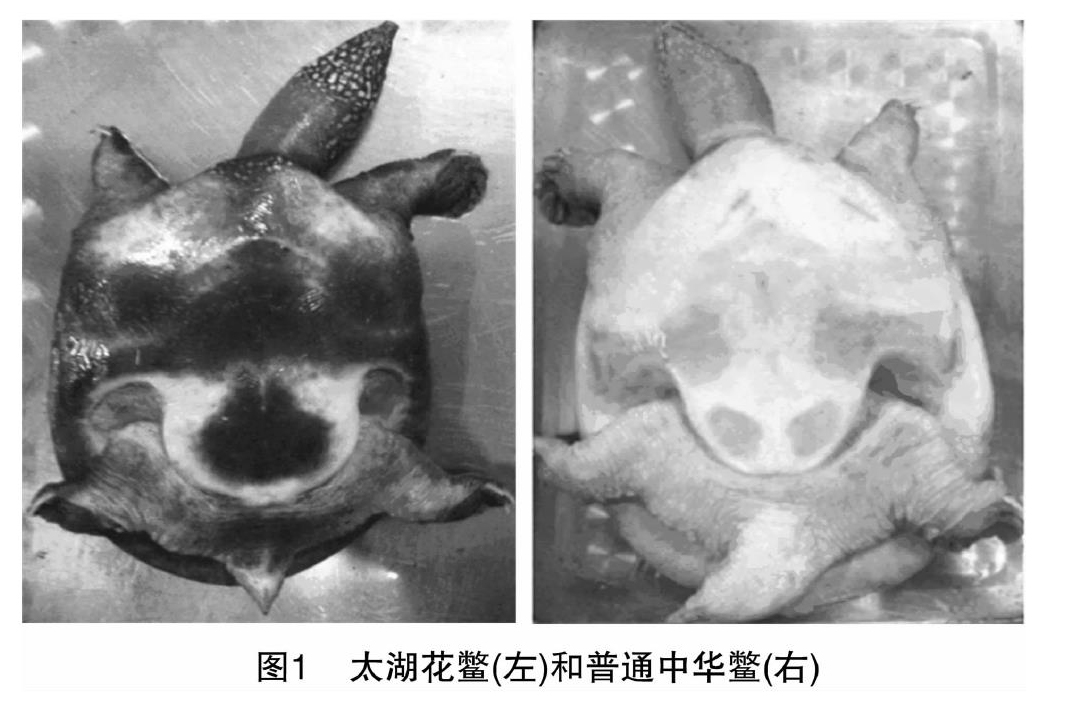
太湖花鳖属于中华鳖(Trionyxsinensis)的地方特色品种,主要分布于长江中下游,太湖流域居多,与普通中华鳖体色差异较大,特别是在自然状态下体色油绿,背部有对称小圆黑斑,腹部有明显块状灰黑花斑,由此得名;并因其较高的营养价值和良好的口感风味受到人们青睐,江南地区多有养殖,市场前景广为看好。太湖花鳖雌雄体在形态规格及生长特性上存在一定差异,本研究通过多元统计分析和线粒体D-loop控制区分析,于表型和基因型多方面对太湖花鳖雌雄特征予以揭示,以期为太湖花鳖雌雄鉴别,以及种质鉴定、资源保护和利用提供一定的理论支持。
1材料与方法
1.1材料来源
太湖花鳖样品是在2017—2018年取自太湖常州武進段的野生群体,共计取样30只,其中,雌雄各半,试验于2019年在长江特色水产工程技术研究中心完成。太湖花鳖活体用于形态参数统计分析,之后取样品腿部肌肉组织于-70℃冰箱中保存,以备提取DNA使用。
[FK(W11][TPWMM11.tif;S+3mm][FK)]
1.2形态参数测量
游标卡尺测量每尾太湖花鳖的体高、头长和头宽等16项形态指标(精度为0.1mm),电子天平测量每尾太湖花鳖的体质量(精度为0.1g),具体测量方法参考国家标准(GB2104—2007)[14],具体指标见表1、表2。鉴于活体太湖花鳖不便于测量,故可将其放入密闭容器,进行乙醚吸入式麻醉后方可精确测量。
1.3D-loop控制区的PCR扩增及测序
参照韩晓磊等的研究方法[15]提取太湖花鳖样品基因组DNA并检测其完整性,测定其浓度后于-20℃保存备用。太湖花鳖2对扩增D-loop控制区的引物序列分别为:T1(5′-3′):[JP9]ATCTTACCCCTACTACACACAT;T2(5′-3′):[JP9]AGACTGGTGATGGAGTATC;T3(5′-3′):[JP9]GCCACAATACTTGTTTATCT;T4(5′-3′):[JP9]AATATCCATCTTGGCGTCTTCA。PCR扩增反应参见陆文浩等的方法[16]具体操作,并测得太湖花鳖样品线粒体DNA序列。
1.3数据处理
1.3.1形态参数分析
试验数据采用SPSS19.0软件进行分析,本研究多元分析方法包括t-检验和主成分分析,它们是针对多个研究对象同时进行分析所得数据的运算方法。通过将背甲长作为基数,以取得2个形态参数比值,从而消除鳖体规格大小对多元分析中参数值的影响。
1.3.2D-loop控制区序列分析
所获得太湖花鳖D-loop控制区序列经过校对处理后,采用软件ClustalX2.1和MEGA5.1进行序列比对和分析,通过Kimura双参数模型统计雌雄群体群间遗传差异,运用邻接法构建系统发生树,以bootstrap进行检验,重复1000次。
2结果与分析
2.1形态差异分析
由表1可知,太湖花鳖雄鳖体质量与尾长关系的r=0.662(r>r0.01),尾部生长与体质量增加相关性显著,其一元线性回归方程为:Y体质量=44.473X尾长+115.837,回归显著性检验F=10.143,F>F0.01;太湖花鳖雌鳖体质量与尾长关系的r=0.445(r 2.2主成分分析 由表2可知,太湖花鳖雌雄群体方差贡献率较高的4个主成分贡献率分别为43.163%、14.427%、8.812%和8.600%,其累积贡献率为75.003%,包含了群体总变异的大部分,可见以上4个相互独立的主成分可代表太湖花鳖雌雄群体间的形态差异。在主成分1中,变量X2、X3、X4、X6、X7、X8、X9、X12和X13影响最大;在主成分2中,变量X10、X11和X14影响最大;在主成分3和主成分4中,变量X2影响最大。背甲周长、背甲宽、腹甲长、腹甲宽和尾长是主成分1变异贡献的主要形态指标,可见太湖花鳖鳖体长度、宽度和尾长是引起其雌雄群体形态差异的主要指标。 由图2可知,太湖花鳖雌雄群体主成分1和主成分2为相对值,雌雄群体之间存在部分重叠,主成分1和主成分2不能准确将雌雄群体进行区分,故太湖花鳖雌雄群体总体形态差别较小,形态差异统计分析较难将其准确鉴别。 2.3D-loop控制区序列分析 2.3.1太湖花鳖线粒体D-loop控制区序列比较 测序所得序列与GenBank中序列号为AY687385.1的中华鳖线粒体DNA序列进行Blast同源性比对,证实所测序列位于第15321位到15835位之间,大小为515bp,碱基组成分别为G、C、T和A的碱基平均含量分别为10.0%、30.0%、32.9%和27.1%,其中T+A含量(60.0%)高于G+C含量(40.0%),是线粒体DNA典型的反G偏倚特征。太湖花鳖雌雄群体的群内遗传相似度为99.23%。 2.3.2太湖花鳖分类地位鉴定 根据D-loop控制区序列差异,用MGEA5.1分析2个群体的遗传距离,由图3可知,发现雌雄群体间遗传差异为0.0114。构建聚类系统发生树,太湖花鳖雌雄群体混为一体,群内个体无单独聚类。 3讨论 t检验是用于小样本(样本容量小于30)的2个平均值差异程度的检验方法,它是用T分布理论来推断差异发生的概率,从而判定2个平均数的差异是否显著[17-18]。本研究中,太湖花鳖雄性尾巴的生长与其体质量的增加显著相关,可用于太湖花鳖雌雄性别的鉴别;主成分分析中尾长也是引起形态差异的主成分变量之一,且需与其他形态变量共同作用进行差异分析。珠水对中华鳖性别鉴别方法进行了研究,指出雄性中华鳖较之雌性尾巴较长,大多能自然伸出裙边外,故尾部特征可以作为雌雄判别的重要依据[19],此结论与本研究结果基本一致。然而,在太湖花鳖雌雄群体30个样本的散点图分析结果中,太湖花鳖雌雄群体并没有明显区分。以上推斷表明,太湖花鳖个体形态差异与其性别存在一定程度的相关性,特别是尾部特征,可用于雌雄性别的初步判断,但其群体的形态差异并不明显,还需更多的参数乃至更深入的分析研究予以揭示。 本研究通过线粒体D-loop控制区序列比对差异分析,发现太湖花鳖雌雄群体群间遗传差异仅为0.0114,可以认为2个群体间不存在有效遗传差异。太湖花鳖雌雄群体聚类系统发生树中2个群体的群内个体未按照性别差异单独聚类,同样揭示雌雄群体群间遗传差异并不明显。 综上所述,太湖花鳖雌雄群体在外观形态上可通过尾巴长短进行简单区分,但进行准确的分辨还相对较难,线粒体DNA差异同样不明显。由此推断,太湖花鳖雌雄群体差异在外观形态只有微小的体现,而在水产养殖中,雄鳖生长速度快于雌鳖,成熟的雄性个体比雌性大且一般为雌性的3倍左右[17],揭示太湖花鳖雌雄差异可能主要体现在生长速度上,故体质量的变化可作为研究方向。 参考文献: [1]王立宾.多元统计分析:模型、案例及SPSS应用[M].北京:经济科学出版社,2010:89-91. [2]GolubtsovAS,BerendzenPB.Morphologicalevidencefortheoccurrenceoftwoelectriccatfish(Matapterurus)speciesintheWhiteNileandOmo-Turkanasystems(EastAfrica)[J].JFishBio,1999,55(3):492-505. [3]GibsonAR,BakerAJ,MoeedP.Morphometricvariationinintroducedpopulationsofthecommonmyna(Acridotherestristis):Anapplicationofthejaekknifetopfincipalcomponentanalysis[J].SystematicZoology,1984,33(4):408-421. [4]VossRS,MarcusLF,PatriciaEP.MorphologicalevolutioninmuroidrodentsⅠ.conservativepatternsofcraniometriccovarianceandtheirontogeneticbasisintheneotropicalgenuszygodontomys[J].Evolution;InternationalJournalofOrganicEvolution,1990,44(6):1568-1587. [5]SomersKM.Multivariateallometryandremovealofsizewithprincipalcomponentsanalysis[J].SystematicZoology,1986,35(3):359-368. [6]潘德博,陈昆慈,朱新平,等.黄喉拟水龟(♀)与三线闭壳龟([XZ(20#]♂[XZ)])杂交后代的形态特征及其与父母本的比较研究[J].水生生物学报,2009,33(4):620-626. [7]陈大庆,张春霖,鲁成,等.青海湖裸鲤繁殖群体线粒体基因组D-loop区序列多态性[J].中国水产科学,2006,13(5):800-806. [8]肖武汉,张亚平.鱼类线粒体DNA的遗传与进化[J].水生生物学报,2000,24(4):384-391. [9]谭围.孟加拉笛鲷和四带笛鲷线粒体基因组特征及分子进化研究[D].湛江:广东海洋大学,2009. [10]董晓丽.半滑舌鳎(Cynoglossussemilaevis)线粒体基因组全序列和性别相关基因的克隆与表达分析[D].青岛:中国海洋大学,2010. [11]李胜杰,白俊杰,叶星,等.基于线粒体D-loop区探讨我国养殖大口黑鲈的分类地位和遗传变异[J].海洋渔业,2008,30(4):291-296. [12]盛军庆,林巧惠,王军花,等.池蝶蚌线粒体基因组双单性遗传现象分析[J].动物学杂志,2014,49(4):597-604. [13]薛婷.淡水珍珠蚌DUI发生及性腺发育研究[D].上海:上海海洋大学,2016. [14]张轩杰,刘筠,周工健,等.中华鳖国家标准:GB21044—2007[S].北京:中国标准出版社,2007. [15]韩晓磊,徐建荣,李小蕊,等.鳡鱼群体遗传多样性的AFLP分析[J].南京师大学报(自然科学版),2009,32(1):110-114. [16]陆文浩,韩晓磊,陈梁,等.不同群体中华鳖的形态多样性分析[J].水产科学,2017,36(6):784-788. [17]秦秋燕.基于动物觅食原理的改进微粒群算法研究[D].太原:太原科技大学,2010. [18]李艳.数据分析软件SAS实用教程[M].武汉:武汉大学出版社,2015:299-301. [19]珠水.中华鳖雌雄鉴别简易方法[J].现代渔业信息,2004,19(11):33.
- 解读当代女性艺术家笔下的女性形象
- 论《一个人的战争》的女性主义特征
- 浅析男权意识笼罩下《月牙儿》的女性悲歌
- 试运用“青蛙效应”来克服职场人员的现代性悲剧
- 浅析《金锁记》
- 《边城》悲剧意蕴探析
- 试论史铁生创作中的个人乌托邦想象
- 试述《官场现形记》中芜湖关道刁迈彭形象及其认识意义
- 论郁达夫《迟桂花》中的三组映照式人物关系
- 林黛玉在曹雪芹本红楼梦80回后身份探佚
- 二十世纪初摄影技术在漫画中的应用
- 文化宣传对人们日常生活的影响
- 浅谈社区群众文化建设
- 初探社会力量参与公共文化服务体系建设
- 新时期高校艺术设计专业应用技能型人才的培养
- 浅谈对小学低段学生阅读兴趣的培养
- 农村美术课堂教学中兴趣的培养
- 浅谈音乐教育对感知觉发展的影响
- 创新初中音乐教学
- 服装设计教学中对学生品牌服装设计思维能力的培养探讨
- 中学音乐欣赏课教学之我见
- 浅谈美术基础教学
- 浅析高职院校共青团构建志愿服务体系与措施
- 包装结构与容器造型课程的教学要点概述
- 慕课背景下的艺术设计类课程教学
- assault²
- assault¹
- assed
- assemble
- assembled
- assembles
- assemblies
- assemblies'
- assembling
- assembly
- assemblyline
- assembly line
- assembly lines
- assembly point
- assemblypoint
- assembly's
- assembly worker
- assemblyworker
- assen
- assent
- assentant
- assented
- assenter
- assenters
- assenting
- 弃繻
- 弃繻人
- 弃繻入关
- 弃繻关
- 弃繻生
- 弃约
- 弃绝
- 弃绝事理
- 弃绝尘俗
- 弃绝聪明智巧,力求返璞归真
- 弃置
- 弃置、抛弃
- 弃置不用
- 弃置不用的优秀人才和其他好东西
- 弃置无用之物
- 弃置未用的人才
- 弃置未用的贤才
- 弃置的磨石
- 弃置闲散
- 弃置,舍弃
- 弃群臣
- 弃考
- 弃职
- 弃职,辞官
- 弃背