姜竹 陈丹丹 刘宏波


摘要:花生(Arachis hypogaea)是我国的主要油料作物,花生油脂的合成受3-磷酸甘油酰基转移酶(GPAT)调控,但其调控机制尚不清楚。从花生中克隆了13个GPAT基因,编码含有4个酰基转移酶保守结构域的蛋白;同时,运用酵母遗传互补体系对基因功能进行了鉴定。在该体系中,酵母双突变体菌株ZAFU1(Δgat1/Δgat2)因缺少GPAT而不能在特定的葡萄糖培养基上生长。结果表明,AhGPAT1/2/3/6/9与空白载体一样,其导入并不能恢复酵母双突变体ZAFU1在葡萄糖中的生长缺陷,而AhGPAT5B、AhGPAT7A/B的导入能恢复酵母双突变体ZAFU1在葡萄糖中的生长缺陷,说明上述3个基因具有酰基转移酶活性,从而为进一步研究花生油脂的合成代谢调控提供了参考。
关键词:花生;3-磷酸甘油酰基转移酶(GPAT);遗传互补;酵母双突变体菌株
中图分类号: S565.201 ?文献标志码: A ?文章编号:1002-1302(2019)22-0056-05
油脂是高等植物的主要能量储存形式,可为植物的种子萌发、花粉发育和有性生殖等提供能量,对于植物的生长发育至关重要[1]。植物油可为人类提供能量和营养,同时也是重要的工业原料[2]。我国的花生(Arachis hypogaea)总产量、单位面积产量和出口量一直位居我国油料作物之首,在我国油脂供应、出口创税和国民经济发展中占有举足轻重的地位,但是花生油脂的合成机制目前尚不清楚[3-4]。
植物油的主要成分是三酰甘油(TAG),由3-磷酸甘油酰基转移酶(GPAT)、溶血磷脂酸酰基转移酶(LPAAT)和二酰甘油酰基转移酶(DGAT)依次催化脂肪酸链掺入甘油骨架的sn-1、sn-2、sn-3位而生成[5-6]。目前,关于甘油脂合成前2步酰化反应的酶,特别是第1步酰化反应的酶知之甚少。研究者在拟南芥、油菜、水稻和向日葵等多种植物中已成功克隆了多个编码酰基转移酶的GPAT基因[7],但普遍认为,除了ATS1,只有GPAT9直接参与甘油脂的合成,其他GPAT是否参与甘油脂合成并不清楚[8-10]。已有研究者从花生中克隆了部分GPAT基因,并对其表达特性进行了分析,但是关于花生GPAT酶的特性尚无相关报道[11-12]。本研究从山花15中分离了13个3-磷酸甘油酰基转移酶基因,并用酵母遗传互补体系对上述基因进行了功能鉴定,发现AhGPAT1/2/3/6/9不能恢复酵母双突变体ZAFU1在葡萄糖中的生长,AhGPAT5B、AhGPAT7A/B能恢复酵母双突变体ZAFU1在葡萄糖中的生长,说明上述3个基因具有酰基转移酶活性。
1 材料与方法
1.1 试验材料
花生品种山花15、酵母菌株ZAFU1[13]、大肠杆菌菌株DH5α和酵母表达载体pYES2-yADH1-Kan V2[14]均由笔者所在实验室保存。RNA提取试剂盒,购自杭州新景生物试剂开发有限公司;质粒提取试剂盒、PCR产物回收试剂盒、胶回收试剂盒,购自生工生物工程(上海)股份有限公司;高保真酶PrimeSTAR、PrimeScriptTM Ⅱ 1st Strand cDNA Synthesis Kit反转录试剂盒、DNA连接酶以及限制性内切酶,购自TaKaRa公司。引物由赛默飞世尔科技有限公司合成。基因测序由铂尚生物技术有限公司完成。
1.2 RNA的提取及cDNA的合成
参照植物总RNA提取试剂盒说明书提取花生根、叶的总RNA。测定总RNA的浓度和纯度,用琼脂糖凝胶电泳检测验证RNA的完整性,用反转录试剂盒反转录合成cDNA。
1.3 基因克隆和重组载体的构建
参考已有的研究,检索得到花生GPAT1/2/3/5/6/8/9基因的核苷酸序列,以这些基因结合拟南芥GPAT氨基酸序列,在花生数据库peanutbase中进行同源检索,得到候选AhGPAT序列,基于此确定花生GPAT1A/1B/2A/2B/3B/5B/6A/7A/7B/8A/9A-1/9A-2/9B序列的信息。结合候选基因序列和表达载体酶切位点,在SIGMA网站上设计各基因的PCR扩增引物(表1)。
以花生cDNA为模板,用带有酶切位点的引物进行RT-PCR扩增,并对目的片段进行胶回收。将酶切后的AhGPAT与酵母表达载体pYES2-yADH1-Kan V2进行连接,转化至大肠杆菌DH5α中,涂布在含有50 mg/L卡那霉素的LB培养基上,37 ℃过夜培养,次日进行菌落PCR检测,并进行测序和序列比对。最后提取测序成功的AhGPAT-pYES2-yADH1-Kan V2重组质粒备用。
1.4 系统发育树分析
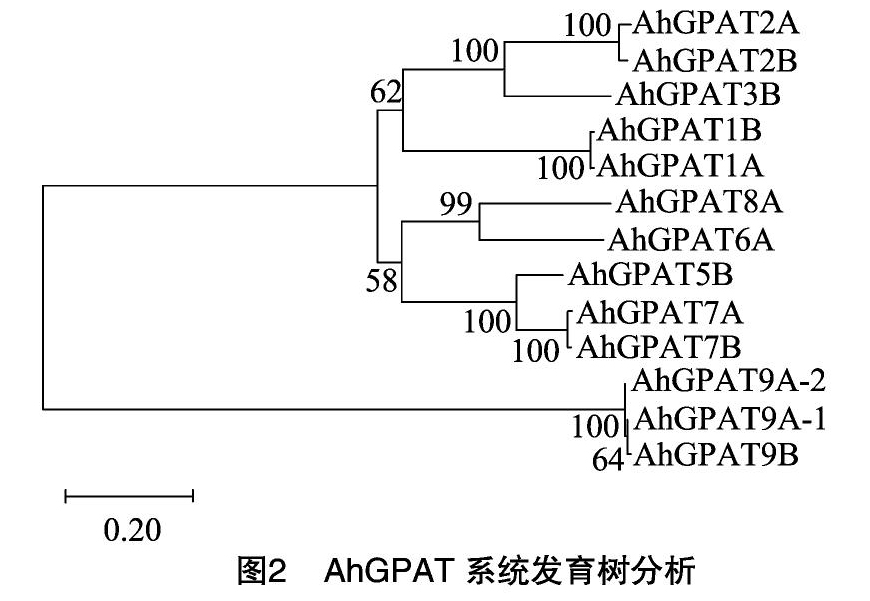
使用DNAMAN对已克隆的10个AhGPAT基因(AhGPAT1A/1B/2A/2B/3B/5B/6A/7A/7B/8A)编码的蛋白氨基酸序列进行比对,使用MEGAX软件对AhGPAT的氨基酸序列进行系统发育树分析,系统发育树采用Neighbour-Joining(邻接)方法[15-16]构建。
1.5 AhGPAT-pYES2-yADH1-Kan V2载体转化酵母双突变体及酵母浓度梯度稀释试验
通过LiAc法制备ZAFU1酵母感受态细胞,将重组质粒AhGPAT-pYES2-yADH1-Kan V2、pYES2-yADH1-Kan V2空白载体和GAT1-pYES2-yADH1-Kan V2陽性对照转入酵母双突变体ZAFU1感受态中。分别涂布于含有葡萄糖(glucose)、半乳糖(galactose)的SC-Ura-His-Leu固体培养基上,于30 ℃培养4~7 d。挑选SC-Ura-His-Leu+葡萄糖固体培养基中的酵母单菌落进行摇菌培养,将菌液分别稀释至D600 nm=1、0.2,0.04、0.008、0.000 16,取5 μL分别涂布于SC-Ura-His-Leu+半乳糖、SC-Ura-His-Leu+葡萄糖固体培养基上,于30 ℃培养4~7 d。
2 结果与分析
2.1 不同AhGPAT氨基酸序列的差异分析
本研究分离了AhGPAT的8个不同成员,共13个基因。分离的由AhGPAT编码的蛋白羧基端均有典型的酰基转移酶的4个保守结构域(AhGPAT9除外):AT-Ⅰ、AT-Ⅱ、AT-Ⅲ 和AT-Ⅳ(图1),保守结构域中与酰基转移酶催化活性相关的氨基酸是AT-Ⅰ 的组氨酸(His)及天冬氨酸(Asp)、AT-Ⅲ 的甘氨酸(Gly)、AT-IV的脯氨酸(Pro);与底物甘油绑定相关的氨基酸是AT-Ⅱ的精氨酸(Arg)、AT-Ⅲ 的谷氨酸(Glu)。
为了更好地确定花生GPAT的亲缘关系,对AhGPAT进行系统发育树分析,并对部分AhGPAT进行氨基酸序列比对。
结果显示,分离的由AhGPAT编码的蛋白之间的氨基酸序列相似度为19.1%~99.7%。栽培花生A/B 2个亚基因组和亚基因组内由AhGPAT编码的蛋白氨基酸相似度极高,为 96.3%~99.7%。系统发育树结果表明,分离的由AhGPAT编码的蛋白可聚成两大分支:AhGPAT1/2/3/5/6/7/8和AhGPAT9(图2)。其中,AhGPAT1/2/3/5/6/7/8蛋白可分为3个亚家族:AhGPAT1/2/3、AhGPAT5/7和AhGPAT6/8;AhGPAT2和AhGPAT3的亲缘关系较近,AhGPAT3B氨基酸序列与AhGPAT2A、AhGPAT2B氨基酸序列的相似度分别为73.0%、73.2%。AhGPAT5/7的亲缘关系较近,AhGPAT5B氨基酸序列与AhGPAT7A、AhGPAT7B氨基酸序列的相似度分别为80.4%、80.9%。AhGPAT6A与AhGPAT8A的亲缘关系较近,二者的氨基酸序列相似度为74.9%。AhGPAT9与其他AhGPAT的亲缘关系较远。
2.2 AhGPAT的功能分析
2.2.1 重组酵母表达载体的构建 对AhGPAT-pYES2-yADH1-Kan V2重组质粒进行酶切鉴定,获得如图3所示与预期大小一致的目的条带,表明AhGPAT-pYES2-yADH1-KanV2重组质粒已构建成功。
2.2.2 酵母遗传互补及酵母浓度梯度稀释试验 笔者所在实验室构建了GAT1、GAT2双敲除的ZAFU1(gat1Δgat2Δ+[pGAL1::At1g06520 Leu2])菌株,其本身含有His3筛选标记, 同时含有Leu2筛选标记的YEplac181-AtGPAT1重组质粒,该质粒GAL1启动子在半乳糖条件下能够诱导AtGPAT1基因的表达,但在葡萄糖条件下,AtGPAT1基因不能表达。因此该酵母双突变体ZAFU1只能在SC+Ura-His-Leu+Galactose培养基中生长,不能在SC+Ura-His-Leu+Glucose培养基中生长。pYES2-yADH1-Kan V2空白载体含有合成尿嘧啶的基因,该质粒yADH1启动子受葡萄糖诱导,受半乳糖抑制,因此转入该载体的酵母菌株若能在 SC-Ura-His-Leu+Glucose 培养基上生长,则证明转入酵母双突变体ZAFU1的目的基因具有酰基转移酶活性。
本研究将转化重组质粒AhGPAT-pYES2-yADH1-Kan V2、pYES2-yADH1-Kan V2空白载体和GAT1-pYES2-yADH1-Kan V2的酵母双突变体分别培养在SC-Ura-His-Leu+Galactose、SC-Ura-His-Leu+Glucose培养基上,发现转pYES2-yADH1-Kan V2空白载体和AhGPAT1A/1B/2A/2B/3B/6A/9A-1/9A-2/9B的酵母双突变体均不能在SC-Ura-His-Leu+Glucose培养基上生长,而转GAT1阳性对照、AhGPAT5B、AhGPAT7A/B的酵母双突变体能在SC-Ura-His-Leu+Glucose培养基上生长,说明AhGPAT5B、AhGPAT7A/B具有酰基转移酶活性(图4)。将转AhGPAT5B、AhGPAT7A/B的酵母双突变体进行浓度梯度稀释培养后,发现它们均能很好地恢复酵母双突变体ZAFU1在葡萄糖培养基上的生长(图5)。转AhGPAT8A的ZAFU1酵母在SC-Ura-His-Leu+Glucose上有少数几个酵母菌落生长,其功能有待进一步探究。将AhGPAT转入条件致死型酵母双突变体ZAFU1中,从葡萄糖或半乳糖培养基上挑取单个菌落稀释(1 ∶ 5)后,分别涂布在SC-Ura-His-Leu+Glucose、SC-Ura-His-Leu+Galactose固体培养基上。
3 结论与讨论
本研究从花生中分离了13个3-磷酸甘油酰基转移酶基因,编码的蛋白均具有酰基转移酶的4个保守结构域[17]。运用酵母遗传互补体系对AhGPAT的酰基转移酶活性进行了鉴定,发现AhGPAT5B/7A/7B能恢复酵母双突变体ZAFU1在葡萄糖培养基中的生长,表明其具有酰基转移酶活性。
拟南芥AtGPAT1具有酰基转移酶活性,而花生AhGPAT1A、AhGPAT1B在酵母遗传互补体系中均不能恢复酵母双突变体ZAFU1的生长,AhGPAT1A/B与AtGPAT1的氨基酸相似度分别只有65.8%、66.1%,AhGPAT1A/B与AtGPAT1的氨基酸序列差异较大,目前尚不清楚这些序列差异是否影响了AhGPAT1A/B的酰基转移酶活性。
根据前人的研究及花生基因组数据库信息,AhGPAT5、AhGPAT7在根、花中的表达量较高,但是这些基因在种子中的表达量较低,而花生根和花的油脂含量低[11]。目前尚不清楚这些基因是否参与种子中油脂、膜脂合成,AhGPAT5、AhGPAT7的功能差異有待进一步探究。
参考文献:
[1]dos Santos Maraschin F,Kulcheski F,Segatto A,et al. Enzymes of glycerol-3-phosphate pathway in triacylglycerol synthesis in plants:function,biotechnological application and evolution[J]. Progress in Lipid Research,2019,73:46-64.
[2]Durrett T P,Benning C,Ohlrogge J B. Plant triacylglycerols as feed stocks for the production of biofuels[J]. The Plant Journal,2008,54(4):593-607.
[3]万书波. 我国花生产业面临的机遇与科技发展战略[J]. 中国农业科技导报,2009,11(1):7-12.
[4]许婷婷,宫清轩,江 晨,等. 我国花生产业的发展现状与前景展望[J]. 山东农业科学,2010(7):117-119.
[5]Li-Beisson Y H,Shorrosh B,Beisson F,et al. Acyl-lipid metabolism[J]. Arabidopsis Book,2013(11):e0161
[6]Wang L P,Shen W Y,Kazachkov M,et al. Metabolic interactions between the Lands cycle and the Kennedy pathway of glycerolipid synthesis in Arabidopsis developing seeds[J]. The Plant Cell,2012,24(11):4652-4669.
[7]Waschburger E,Kulcheski F R,Veto N M,et al. Genome-wide analysis of the glycerol-3-phosphate acyltransferase (GPAT) gene family reveals the evolution and diversification of plant GPATs[J]. Genetics and Molecular Biology,2018,41(suppl 1):355-370.
[8]Shockey J,Regmi A,Cotton K,et al. Identification of Arabidopsis GPAT9 (At5g60620) as an essential gene involved in triacylglycerol biosynthesis[J]. Plant Physiology,2016,170(1):163-179.
[9]Kunst L,Browse J,Somerville C. Altered regulation of lipid biosyn thesis in a mutant of Arabidopsis deficient in chloroplast glycerol-3-phosphate acyltransferase activity[J]. Proceedings of the National Academy of Sciences of the United States of America,1988,85(12):4143-4147.
[10]Singer S D,Chen G,Mietkiewska E,et al. Arabidopsis GPAT9 contributes to synthesis of intracellular glycerolipids but not surface lipids[J]. Journal of Experimental Botany,2016,67(15):4627-4638.
[11]郝翠翠,梁成偉,石 蕾,等. 花生甘油-3-磷酸酰基转移酶(GPAT)基因的克隆及表达分析[J]. 花生学报,2018,47(1):1-10,18.
[12]Chi X Y,Yang Q,Pan L,et al. Isolation and expression analysis of glycerol-3-phosphate acyltransferase genes from peanuts(Arachis hypogaea L.)[J]. Grasasy Aceites,2015,66(3):e093.
[13]Lei J,Miao Y C,Lan Y,et al. A novel complementation assay for quick and specific screen of genes encoding glycerol-3-phosphate acyltransferases[J]. Frontiers in Plant Science,2018,9:353.
[14]陈丹丹,刘宏波. 筛选GPAT基因的酵母遗传互补体系的优化[J]. 江苏农业科学,2019,47(13):64-66.
[15]李向阳,汤 宏,张国辉. 比较基因组分析斯氏假单胞菌遗传多样性及固氮基因岛进化机制[J]. 江苏农业科学,2017,45(1):33-38.
[16]禹海鑫,安榆林,郭骁驹,等. 不同种类白条天牛基因条形码特征与系统发育初探[J]. 江苏农业科学,2017,45(13):90-94.
[17]Lewin T M,Wang P,Coleman R A. Analysis of amino acid motifs diagnostic for the sn-glycerol-3-phosphate acyltransferase reaction[J]. Biochemistry,1999,38(18):5764-5771.
- 有效提问,生成精彩数学课堂
- 体“味”——有数学味的数学课
- 有效提问让课堂思维更深刻
- 曲径通幽之“几何直观”微探
- 用阶段性体验,催生数学思想的花朵
- 让教学从学生出发
- 巧借导入艺术,优化小学数学课堂教学
- 预习之后的数学课堂教学实施策略
- 例谈如何在“过程”中引领学生思考
- 实现“四化”,构建高效课堂
- 例谈“以退为进”策略在数学学习中的运用
- 关于低年级童话式情境导学策略的几点思考
- 浅谈数学课堂教学中教师的“导”
- “三多三少”,促进学生的可持续发展
- 运用概念图绘制概念教学的思维蓝图
- 从细处入手打造美丽课堂
- 用游戏展现数学之美
- 从实践的角度谈数学教学
- 学生计算,要给思考的支撑点
- 将学生的思维引向深处
- 有效源自真实朴实扎实
- 精彩源于细微处
- 巧用“现场资源” 促进有效生成
- 把握教学要求 凸显知识本质
- 遵从学生的真实思维,实现有效教学
- unjuster
- unjustest
- unjustice
- unjustifiable
- unjustifiableness
- unjustifiablenesses
- unjustifiably
- unjustified
- unjustifiedly
- unjustly
- unjustness
- unjustnesses
- unjuvenile
- unjuvenilely
- unjuvenileness
- unjuvenilenesses
- unkeeled
- unkempt
- unkempter
- unkemptest
- unkemptly
- unkemptness
- unkemptnesses
- unkerchiefed
- unkey
- 仪表风度不同一般
- 仪表风采
- 仪表,多就姿态而言
- 仪观
- 仪贤堂
- 仪质
- 仪车
- 仪轨
- 仪适
- 仪部
- 仪采
- 仪锽
- 仪门
- 仪静体闲
- 仪韶
- 仪马
- 仪鸾司
- 仪鸾司大使
- 仫
- 仫佬族
- 仫佬族的传统节日风俗典故
- 们
- 们浑
- 仰
- 仰不剌叉