谢文 郅军锐 刘利 吕莎莎 周治成

摘要:为明确西花蓟马取食不同嗜食寄主对其体内胰蛋白酶基因CL4520.Contig1表达量的影响,研究了西花蓟马从菜豆豆荚分别转换到较嗜食寄主菜豆植株和非嗜食寄主蚕豆植株上继代饲养1、2、3代后,其2龄若虫和成虫体内CL4520.Contig1表达量的异同。结果表明,转换到菜豆植株上继代饲养的1、2、3代西花蓟马2龄若虫体内CL4520.Contig1表达量分别比对照降低了61.86%、13.16%、19.10%,在成虫体内降低了57.07%、88.03%、12.41%;转换到蚕豆植株上继代饲养的1、2、3代西花蓟马2龄若虫体内CL4520.Contig1表达量分别是对照的2.33倍、6.06倍、4.10倍,成虫则是对照的3.93倍、6.44倍、5.55倍。由此得出,当转换寄主后,西花蓟马体内胰蛋白酶基因CL4520.Contig1会迅速产生响应,其表达量发生明显变化,CL4520.Contig1的表达量在取食嗜食寄主菜豆植株时受到一定的抑制,而在取食非嗜食寄主蚕豆植株时明显被激活,且CL4520.Contig1的表达量在西花蓟马不同龄期存在差异。
关键词:西花蓟马;不同嗜食寄主;胰蛋白酶;基因表达量
中图分类号: S433.89;Q786文献标志码: A
文章编号:1002-1302(2020)07-0069-05
寄主植物和昆虫在长期的进化过程中相互影响、相互适应或协同进化,当寄主植株被昆虫取食时会产生多种防御物质[1],而植食性昆虫面对寄主的防御机制,也会在进化过程中形成相应的反防御机制,其中昆虫体内的消化酶就起到了重要作用[2]。昆虫利用其体内消化酶降解植物淀粉和蛋白质等,达到消化和吸收营养物质的目的;同时,昆虫体内消化酶活性的高低反映了昆虫对营养物质消化吸收的能力和生长发育的速率[3-5]。胰蛋白酶(trypsin,TRY)是昆虫中肠中重要的蛋白水解酶,可以有力迅速地激活其他蛋白酶原,进而行使消化功能[6]。此外,胰蛋白酶还具有阻止前体细胞抗体毁灭的功能[7]。
西花蓟马[Frankliniella occidentalis (Pergande)]属缨翅目(Thysanoptera)蓟马科(Thripidae)。21世纪初,西花蓟马在我国北京首次被报道[8],之后迅速传播扩散,近年来相继在吉林[9]、新疆[10]、宁夏[11]和内蒙古中西部[12]等地被发现。西花蓟马主要通过锉吸式口器危害寄主植物的叶、花和果实[13],也可传播番茄斑萎病毒(tomato spotted wilt virus,TSWV)等多种植物病毒,加重对植物的危害[14-15],是世界危险性害虫。
西花蓟马是典型的多食性害虫,可危害不同种类植物,主要危害花卉、蔬菜和果树等,寄主达60多个科500多种植物,可在不同寄主植物间迁移扩散危害[16]。但西花蓟马在不同寄主植株上的发育历期、产卵量和着虫率等有明显差异[17-19]。由于不同寄主植株所含营养物质的质和量不同[20],昆虫取食诱导产生的防御物质也不同,因此导致昆虫在不同寄主上适应性不同。Kotkar等的研究结果表明,昆虫取食不同寄主植株后消化酶活性都会不同[21-22],说明不同寄主是影响昆虫体内消化酶活性的重要因子。牟峰的研究表明,取食不同处理菜豆植株后,西花蓟马体内消化酶活性发生显著变化[23];姜丽娜等研究发现,取食不同寄主植株的西花蓟马体内消化酶活性变化不一致[6];在蒲恒浒等的研究中,西花蓟马取食不同诱导处理的番茄植株后,其体内消化酶活性显著高于取食健康的番茄植株[24],但這些研究中并未报道不同嗜食寄主对西花蓟马消化酶基因表达量的影响。
为研究不同嗜食寄主对西花蓟马消化酶基因表达量的影响,以及西花蓟马取食不同寄主后的逐代适应过程,本试验在前人明确菜豆植株为西花蓟马较嗜食寄主和蚕豆植株为非嗜食寄主的基础上[18,25],研究了取食菜豆豆荚的西花蓟马分别转换到较嗜食寄主菜豆植株和非嗜食寄主蚕豆植株上继代饲养1、2、3代后,其2龄若虫和成虫体内胰蛋白酶基因CL4520.Contig1的表达量,以期揭示西花蓟马对不同嗜食寄主植物的适应性的异同,为明确西花蓟马与寄主之间的互作机制奠定基础。
1 材料与方法
1.1 材料
1.1.1 供试虫源
西花蓟马采自贵州省贵阳市花溪周边蔬菜地,鉴定后在人工气候箱[RXZ型,购自宁波江南仪器厂,温度为(25±1) ℃,相对湿度为(70±5)%,光—暗周期为14 h—10 h]以菜豆豆荚长期继代饲养达20代以上。
1.1.2 供试寄主
菜豆品种选自河北省辛集市盛农种子公司的金束鹿泰国地豆王,于人工气候室(条件同人工气候箱)内培养菜豆植株,每营养钵(直径10 cm,高9 cm)培育2株清洁苗,待菜豆长至15 cm左右、3~4张真叶时,取长势一致的菜豆植株作为供试寄主植物,生长期间不施用任何农药。
蚕豆品种选自康乐县进忠农产品实业开发有限责任公司的临蚕五号,于人工气候室内培养蚕豆植株(条件同上),每营养钵(直径10 cm,高9 cm)培育4株清洁苗,待蚕豆长至15 cm左右、8~10张叶时,取长势一致的蚕豆植株作为供试寄主植物。生长期间不施用任何农药。
1.2 试验方法
1.2.1 菜豆和蚕豆植株上F1代西花蓟马的饲养及2龄若虫和成虫的取样
选取健康、长势一致的菜豆、蚕豆植株各60钵,每个养虫笼中放置12钵寄主植株,取人工气候箱内菜豆豆荚上饲养的已交配3 d的西花蓟马雌成虫分别接到菜豆植株和蚕豆植株上,每钵植株约100头雌成虫,24 h后,剔除所接所有成虫,每天观察叶片上孵化出的若虫情况,待西花蓟马若虫达2龄阶段,每个养虫笼中挑取200头若虫为1个样,放入1.5 mL离心管中,贴上标签,立即用液氮冷冻于-80 ℃下冰箱中保存,以备提取总RNA时取用。同时保留一定若虫继续在植株上饲养,至成虫阶段后取成虫150头放入1.5 mL离心管中,贴上标签,立即用液氮冷冻于-80 ℃下冰箱中保存,以备提取总RNA时取用。在饲养过程中,如有萎蔫植株,及时补加新鲜长势相同的植株。试验设置5个养虫笼,每个养虫笼的西花蓟马为1个重复。
1.2.2 菜豆和蚕豆植株上F2代西花蓟马的饲养及2龄若虫和成虫的取样
在“1.2.1”节中成虫取样2~3 d后,另取已交配的F1代雌成虫100头,依照“1.2.1”节中相同的方法转移到新的养虫笼健康的同种植株上,待分别发育到2龄若虫和成虫后,对2龄若虫和成虫取样,贴上标签后,立即用液氮冷冻置于-80 ℃下冰箱中保存。试验设置5个养虫笼,每个养虫笼的西花蓟马为1个重复。
1.2.3 菜豆和蚕豆植株上F3代西花蓟马的饲养及2龄若虫和成虫的取样
在“1.2.2”节的中成虫取样2~3 d后,另取已交配的F2代雌成虫100头,依照“1.2.1”节中相同的方法转移到新的养虫笼健康的同种植株上,待分别发育到2龄若虫和成虫后,对2龄若虫和成虫取样,贴上标签后,立即用液氮冷冻置于-80 ℃下冰箱中保存。试验设置5个养虫笼,每个养虫笼的西花蓟马为1个重复。
试验以菜豆豆荚上长期继代饲养的西花蓟马2龄若虫及雌成虫为对照。
1.2.4 西花蓟马成虫和2龄若虫总RNA的提取
参照SV Total RNA分离纯化试剂盒(Promega)说明书步驟进行西花蓟马成虫和2龄若虫总RNA的提取,取吸光度比D260 nm/D280 nm处于1.9~2.1之间及D260 nm/D230 nm为2.1左右的RNA备用。
1.2.5 反转录
根据RevertAid First Strant cDNA Synthesis Kit反转录试剂盒(Thermo)的操作方法进行反转录。
1.2.6 引物设计
引物序列CL4520.Contig1_All-F:CGGTTGTGGAATGGCTAA;CL4520.Contig1_All-R:ACTAGAACGGACGGACTC。引物由生工生物工程(上海)股份有限公司合成。
1.2.7 实时荧光定量(realtime qPCR)分析
以西花蓟马EF-1基因作为内参基因[26]。本试验借助Mx3000PTM荧光定量PCR仪和MxPro软件,以FastStart Essential DNA Green Master(Roche公司)为荧光染料,根据qPCR结果得到的CT值采用2-ΔΔCT法进行基因相对表达量的计算。
1.3 数据分析
用Excel 2016和SPSS 19.0软件对试验数据进行统计和分析,不同世代间差异性分析用Duncans多重比较,2龄若虫和成虫两虫态间、菜豆和蚕豆植株间的差异均采用独立样本t检验。
2 结果与分析
2.1 取食菜豆植株对西花蓟马不同世代胰蛋白酶基因CL4520.Contig1表达量的影响
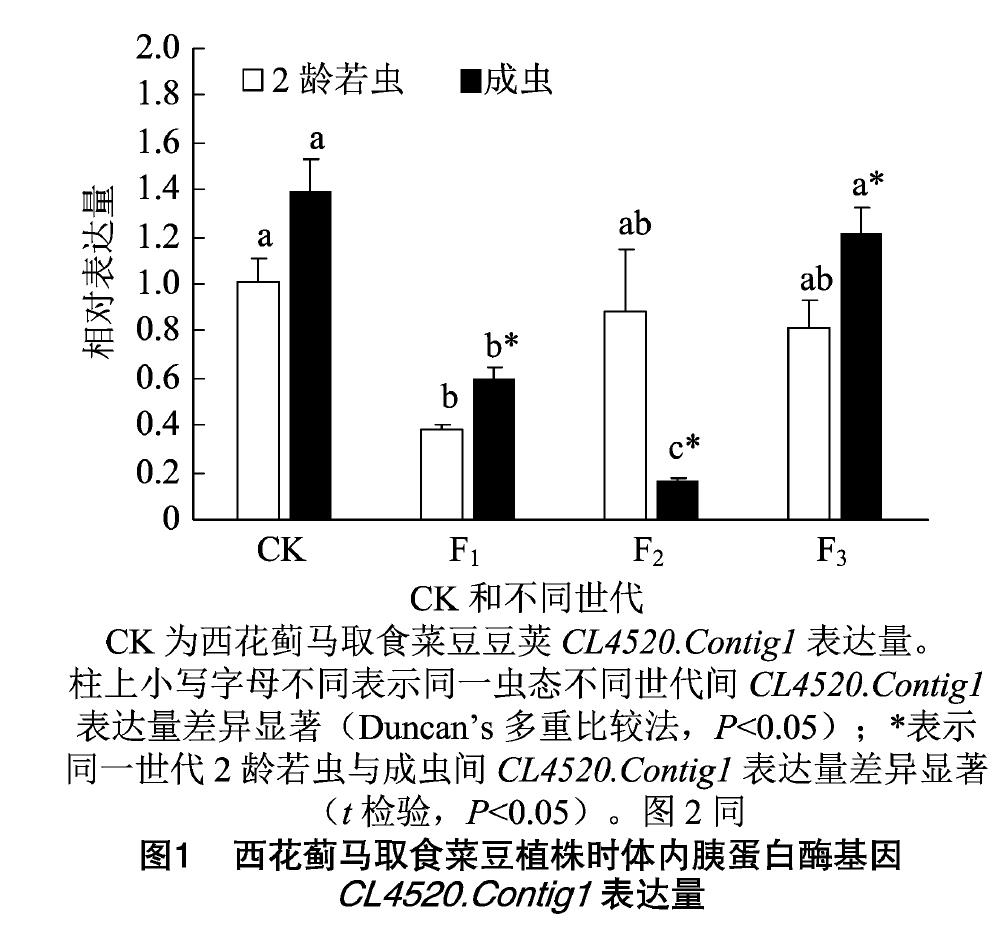
由图1可知,西花蓟马从菜豆豆荚转换到菜豆植株后,西花蓟马2龄若虫体内胰蛋白酶基因CL4520.Contig1的表达量在F1代显著下降;到F2代后,表达量有所上升,与对照和F1代差异均不显著;到F3代CL4520.Contig1的表达量与F2代大致相当。西花蓟马2龄若虫体内胰蛋白酶基因CL4520.Contig1的表达量在F1代、F2代和F3代依次比对照降低61.86%、13.16%和19.10%。
成虫体内胰蛋白酶基因CL4520.Contig1的表达量在F1代也显著下降;到了F2代下降更为明显,达到最低水平,比对照降低了88.03%;到F3代后,西花蓟马成虫体内CL4520.Contig1的表达量上升到了对照水平。
西花蓟马取食菜豆植株F2代时,2龄若虫体内CL4520.Contig1的表达量显著高于成虫;取食菜豆植株F1代和F3代时,均是成虫体内CL4520.Contig1的表达量显著高于2龄若虫。
[FK(W14][TPXW11.tif;S+3mm][FK)]
2.2 取食蚕豆植株对西花蓟马不同世代胰蛋白酶基因CL4520.Contig1表达量的影响
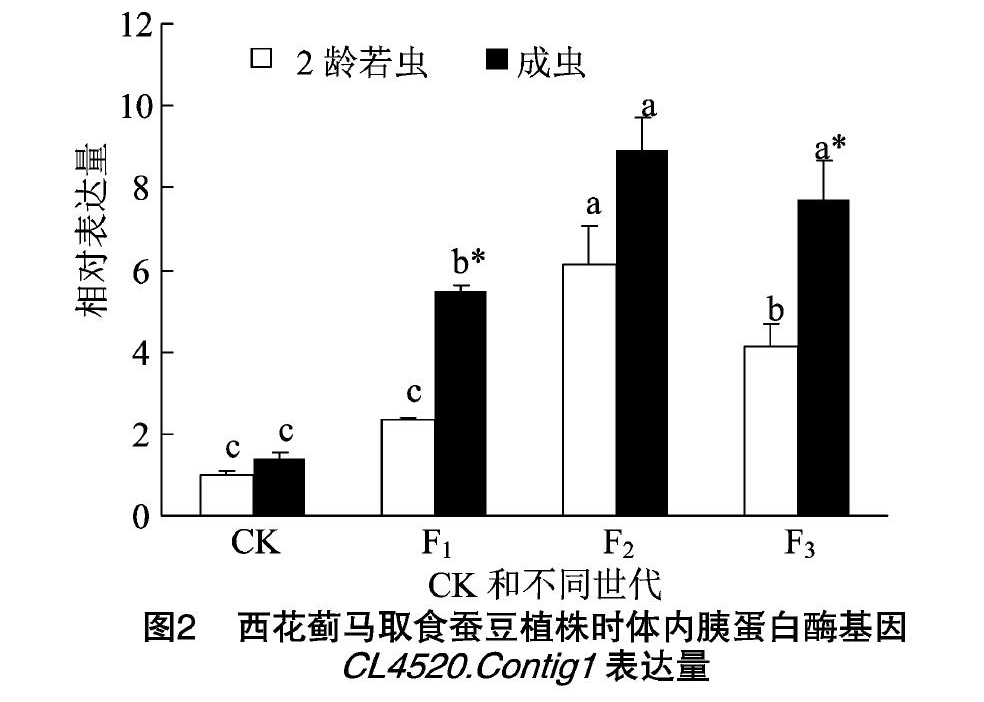
由图2可知,西花蓟马从菜豆豆荚转换到蚕豆植株后,2龄若虫体内CL4520.Contig1的表达量在F1代有所上升但和对照差异不显著;到F2代后,表达量显著上升,达到最高水平,为对照的6.06倍;到F3代CL4520.Contig1的表达量虽比F2代显著下降,但仍显著高于F1代和对照。
成虫体内CL4520.Contig1的表达量转移到蚕豆植株后均显著升高,在F1代和F2代分别为对照的3.93倍和6.44倍,且在F2代达到最高水平,F3代CL4520.Contig1的表达量虽略有下降,但与F2代差异不显著。
西花蓟马取食蚕豆植株F2代时,2龄若虫和成虫体内CL4520.Contig1的表达量差异不显著,而取食蚕豆植株F1代和F3代时,均是成虫体内CL4520.Contig1的表达量显著高于2龄若虫。
2.3 取食不同寄主对西花蓟马体内胰蛋白酶基因CL4520.Contig1表达量的影响
由表1可知,取食不同嗜食寄主植株西花蓟马相同世代和同一虫态下体内胰蛋白酶基因CL4520.Contig1表达量不同,均是取食蚕豆植株的CL4520.Contig1[JP3]表达量显著高于取食菜豆植株的。西花蓟马2龄若虫体内胰蛋白酶基因CL4520.Contig1表达量在取食蚕豆植株的F1代、F2代、F3代分别是取食菜豆植株的F1代、F2代、F3代的6.10倍、6.98、5.07 倍,成虫体内的表达量也是在取食蚕豆植株时高,分别是取食菜豆的9.16、53.82、6.34倍。
3 讨论
昆虫体内的消化酶与取食的食物密切相关,其酶活性的高低是判断昆虫对不同寄主适合度的重要指标[27]。通常情况下,基因表达量上升会导致其编码的蛋白质表达量升高,进而会使其相应酶活性增强[28-29]。本研究发现同一世代和相同虫态下取食非嗜食寄主蚕豆植株的西花蓟马体内胰蛋白酶基因CL4520.Contig1表达量显著高于取食较嗜食寄主菜豆植株的。可能是因为西花蓟马取食非嗜食寄主蚕豆时,为了更好地分解吸收寄主植物营养物质而增加胰蛋白酶基因的表达量,从而增加体内胰蛋白酶的量。此研究结果和Leo等研究结果一致,当斜纹夜蛾取食不适宜寄主时,通过蛋白半胱氨酸消化酶的过量表达来适应植物[30]。但也有研究表明,昆虫取食不同寄主植物时,其消化酶活性与对寄主的嗜食度基本呈正相关,与本研究结果正好相反。如张娜等研究发现,甜菜夜蛾5龄幼虫取食3种不同适合度寄主植物后,其体内淀粉酶和脂肪酶活性随嗜食度的降低而依次降低[31]。王倩倩等的研究表明,取食藜、大豆和向日葵3种喜食寄主的草地螟幼虫中肠消化酶较取食其他2种非喜食寄主植物的中肠消化酶活性更强[32]。阎雄飞等研究发现,取食适宜寄主复叶槭和漳河柳的光肩星天牛成虫消化道的淀粉酶、果胶酶和胃蛋白酶活性显著高于取食抗性寄主新疆杨和白蜡的这3种消化酶的活性[33]。还有研究表明,胰蛋白酶基因表达量与寄主嗜食度无关。如孙洋等研究测定了绿盲蝽取食8种不同嗜食度寄主后体内一种胰蛋白酶基因的表达量,体内AlSP4基因表达量在嗜食寄主和非嗜食寄主上无规律可言,与寄主的嗜食度无关[34-36]。可能是因为不同学者研究的消化酶基因不同,其参与的分子调节机制也不同,也有可能是因为昆虫种类不同,其适应不同寄主的机制也不同,具体原因还需进一步研究。
本研究结果表明,当转换寄主后,F1代西花蓟马2龄若虫和成虫体内胰蛋白酶基因CL4520.Contig1表达量会发生较大变化,除取食菜豆植株的F2代2龄若虫外,其余均在F2代达到最高水平或最低水平,到F3代时,取食菜豆植株的西花蓟马表达量恢复到对照水平,取食蚕豆植株的表达量仍然显著高于对照。可能是转换到较嗜食寄主后,西花蓟马通过调控自身酶基因表达量,在F3代就适应了寄主,而转换到非嗜食寄主后,适应过程较长,在F3代并不能很好地适應寄主,因此继续通过提高酶基因表达量达到适应寄主的目的。
本研究结果表明,西花蓟马取食同种寄主植株的情况下,除了取食菜豆植株的F2代西花蓟马2龄若虫体内胰蛋白酶基因CL4520.Contig1表达量比成虫高,其余的成虫表达量均比同一世代2龄若虫高,说明不同虫态对寄主的适应能力不同。郅军锐等研究发现,西花蓟马2龄若虫和成虫取食健康和二斑叶螨危害后的菜豆植株后体内保护酶和解毒酶活性的变化和虫态、酶的种类及寄主有关,也说明了在适应寄主过程中酶变化的复杂性[37]。
本研究揭示了西花蓟马体内胰蛋白酶基因CL4520.Contig1对菜豆和蚕豆2种不同嗜好寄主的适应过程。至于西花蓟马取食其他不同寄主之后体内胰蛋白酶基因和其他消化酶基因表达量如何改变,不同消化酶基因如何协调,如何适应对不同类别寄主,还须进一步研究。
参考文献:
[1]陈明顺,仵均祥,张国辉. 植物诱导性直接防御[J]. 昆虫知识,2009,46(2):175-186.
[2]彭 露,严 盈,刘万学,等. 植食性昆虫对植物的反防御机制[J]. 昆虫学报,2010,53(5):572-580.
[3]王 琳. 昆虫淀粉酶蛋白质类抑制剂研究进展[J]. 广东农业科学,2006,46(8):110-113.
[4]严 盈,刘万学,万方浩. 唾液成分在刺吸式昆虫与植物关系中的作用[J]. 昆虫学报,2008,51(5):537-544.
[5]周 隆,文礼章,张友军. 不同寄主植物对烟粉虱消化酶和氨基酸代谢酶活性的影响[C]//华中昆虫研究(第七卷). 北京:中国农业科学技术出版社,2011:38-44.
[6]姜丽娜,钱 蕾,喜 超,等. CO2浓度升高对不同寄主植物上西花蓟马和花蓟马成虫体内消化酶活性的影响[J]. 昆虫学报,2017,60(3):237-246.
[7]Ashman R F. Enzymatic modification of lymphocyte receptors for antigen. Ⅲ. Resistance of receptors to trypsin at the peak of the immune response[J]. European Journal of Immunology,1976,5(6):421-425.
[8]张友军,吴青君,徐宝云,等. 危险性外来入侵生物——西花蓟马在北京发生危害[J]. 植物保护,2003,29(4):58-59.
[9]刘若思,刘 燕,王 军,等. 重要外来入侵害虫西花蓟马在吉林省部分地区的首次发现[J]. 北京农学院学报,2015,30(2):24-27.
[10]杨 华,何 伟,崔元玗,等. 西花蓟马在新疆设施蔬菜种植区的发生与分布[J]. 新疆农业科学,2016,53(1):38-42.
[11]张治科,张 烨,吴圣勇. 西花蓟马在宁夏的发生及防控措施[J]. 植物检疫,2016,30(4):75-77.
[12]高振江,张冬梅,高 娃,等. 害虫西花蓟马在内蒙古中西部地区的发生与分布[J]. 北方农业学报,2017,45(2):82-85.
[13]Kirk W J,Terry L I. The spread of the western flower thrips Frankliniella occidentalis (Pergande)[J]. Agric Forest Entomol,2003,5(4):301-310.
[14]Whitfield A E,Ullman D E,German T L. Tospovirus-thrips interactions[J]. Annual Review of Phytopathology,2005,43:459-489.
[15]Adkins S. Tomato spotted wilt virus—Positive steps towards negative success[J]. Molecular Plant Pathology,2000,1(3):151-157.
[16]Manners A G,Dembowski B R,Madaline A H. Biological control of western flower thrips,Frankliniella occidentalis(Pergande)(Thysanoptera:Thripidae),in gerberas,chrysanthemums and roses[J]. Australian Journal of Entomology,2013,52(3):134-137.
[17]李景柱,郅军锐,郑珊珊. 西花蓟马对不同豆科蔬菜寄主的选择性[J]. 贵州农业科学,2010,38(9):103-105.
[18]裴昌莹,郑长英. 西花蓟马对不同蔬菜寄主的选择性研究[J]. 中国生态农业学报,2011,19(2):383-387.
[19]曹 宇,刘 燕,熊正利,等. 西花蓟马对不同花卉寄主的产卵选择性[J]. 植物保护学报,2015,42(5):741-748.
[20]王倩倩,李克斌,尹 姣,等. 不同寄主植物对昆虫生长发育的影响[C]//创新驱动与现代植保——中国植物保护学会第十一次全国会员代表大会暨2013年学术年会论文集.北京:中国农业科学技术出版社,2013.
[21]Kotkar H M,Sarate P J,Tamhane V A,et al. Responses of midgut amylases of Helicoverpa armigera to feeding on various host plants[J]. Journal of Insect Physiology,2009,55(8):663-670.
[22]徐 伟,张吉辉,毕嘉瑞,等. 寄主植物对双斑萤叶甲中肠消化酶和解毒酶活性的影响[J]. 吉林农業大学学报,2018,40(5):23-28.
[23]牟 峰. 不同处理菜豆植株对西花蓟马生长发育和酶活性的影响[D]. 贵阳:贵州大学,2015.
[24]蒲恒浒,郅军锐,曾 广,等. 不同诱导处理对番茄植株保护酶及西花蓟马体内消化酶活性的影响[J]. 植物保护学报,2018,45(4):915-916.
[25]袁成明,郅军锐,曹 宇,等. 西花蓟马对蔬菜寄主的选择性[J]. 生态学报,2011,31(6):1720-1726.
[26]Zheng Y T,Li H B,Lu M X,et al. Evaluation and validation of reference genes for qRT-PCR normalization in Frankliniella occidentalis (Thysanoptera:Thripidae)[J]. PLoS one,2014,9(10):e111369.
[27]贾 冰,谭 瑶,付晓彤,等. 不同寄主植物对牧草盲蝽生长发育、繁殖及消化酶活性的影响[J]. 草业科学,2018,301(8):162-171.
[28]Pan Y O,Guo H L,Gao X W. Carboxylesterase activity,cDNA sequence,and gene expression in malathion susceptible and resistant strains of the cotton aphid,Aphis gossypii[J]. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology,2009,152(3):266-270.
[29]Zhou X J,Sheng C F,Li M,et al. Expression responses of nine cytochrome P450 genes to xenobiotics in the cotton bollworm Helicoverpa armigera[J]. Pesticide Biochemistry and Physiology,2010,97(3):209-213.
[30]Leo F D,Bonadé-Bottino M A. Opposite effects on spodoptera littoralis larvae of high expression level of a trypsin proteinase inhibitor in transgenic plants[J]. Plant Physiology,1998,118(3):997-1004.
[31]张 娜,郭建英,万方浩,等. 寄主植物对甜菜夜蛾生长发育和消化酶活性的影响[J]. 植物保护学报,2009,36(2):146-150.
[32]王倩倩,王 蕾,李克斌,等. 不同寄主植物对草地螟的营养作用及消化酶的影响[J]. 植物保护,2015,41(4):46-51.
[33]阎雄飞,刘 娟,刘永华. 不同寄主对光肩星天牛成虫消化酶的影响[J]. 湖北农业科学,2012,51(18):4010-4013.
[34]徐文华,王瑞明,林付根,等. 棉盲蝽的寄主种类、转移规律、生态分布与寄主的适合度[J]. 江西农业学报,2007,19(12):45-50,56.
[35]魏书艳,肖留斌,谭永安,等. 不同寄主受绿盲蝽危害后生理代谢指标的变化[J]. 植物保护学报,2010,37(4):359-364.
[36]孙 洋,柏立新,张永军,等. 绿盲蝽丝氨酸蛋白酶基因AlSP4的克隆及取食不同寄主植物后的表达谱分析[J]. 昆虫学报,2012,55(6):641-650.
[37]郅军锐,田 甜,温 娟,等. 西花蓟马或二斑叶螨为害的菜豆对两者间后取食者体内保护酶和解毒酶活性的影响[J]. 昆虫学报,2016,59(7):707-715.
- 人本理念在人力资源管理中的应用
- 危险品道路运输优化方案风险评价研究
- 试论广播电视机房的供配电系统设计
- 汕尾市雷电特征及防雷减灾对策
- 房屋建筑施工中防渗漏施工技术要点
- 房屋建筑施工中基坑围护和土方开挖技术应用
- 关于土地管理中遥感技术的应用分析
- 建筑工程中的外墙防渗漏施工技术研究
- 绿色施工管理理念下建筑施工管理创新策略
- 医院护士工作站管理系统的设计与实现
- MES系统中的计划管控技术研究
- 浅析电气自动化技术在电气工程中的应用
- 顶管施工技术在市政工程中的应用分析
- 新时期电力单位如何做好党建工作
- 论程序公正与实体公正的关系
- 试谈20世纪中国社会史的研究
- 以冯玉祥为例看第二次直奉战争前直系内部的矛盾和分裂
- 结合实例探讨未成年女性犯罪的新态势
- 社会堕化的理论发展研究概述
- 持续保持高速公路收费员文明优质服务的思考
- 对曹操性格两面性分析探讨
- 行政事业单位预算绩效管理存在问题与对策
- “师夷长技以制夷”政治思想在旧民主主义革命时期的历史作用
- 浅析英美文学中的人文精神
- 广告的跨文化传播研究
- yellowly
- yellowness
- yellow pages
- yellowpages
- yellow pages™
- yellows
- yellowsheets
- yellow sheets™
- yellowy
- yellow²
- yellow¹
- yells
- yelp
- yelped
- yelping
- yelps
- yen
- yenned
- yenning
- yens
- yep
- yeps
- yes
- yeses
- yes-man
- 性情直爽,心里有话立刻就说出
- 性情直爽,语言明快
- 性情耿直的人
- 性情脾气
- 性情褊急
- 性情豪放,行为散漫
- 性情非常古怪的人
- 性情风度
- 性情高傲
- 性感
- 性擦边
- 性教育
- 性早熟
- 性智
- 性服务
- 性染色体
- 性格
- 性格、心情、神色安静
- 性格、才能)独特而与众不同
- 性格、气质
- 性格、言行古怪,叫人难以捉模
- 性格不同的人并处,或性质不同的事物并存而相安无事
- 性格不好
- 性格不直爽
- 性格不随和,不可接近