刘艳艳 曹婷 刘兴华 严国红

摘要:为明确Pi-ta和Pi9稻瘟病抗性主效基因在32份粳稻骨干亲本中的分布状况,利用抗稻瘟病基因Pi-ta和Pi9的分子标记对32份粳稻骨干亲本进行检测。结果表明,在32份骨干亲本中,Pi-ta抗性基因检出率为 46.88%,Pi9抗性基因检出率为12.50%,同时检测到Pi-ta和Pi9抗病基因,检出率为3.12%,不含有Pi-ta和Pi9抗病基因的材料占比为43.75%。在骨干亲本中,Pi-ta单个基因的检出率较高,Pi9单个基因的检出率较低,同时含有Pi-ta和Pi9抗病基因的材料较少,因此可见,多个抗稻瘟病基因的聚合育种是今后水稻抗病育种的重點。
关键词:水稻;稻瘟病;Pi-ta;Pi9;分子检测;抗性基因
中图分类号: S511.2+20.1;S435.111.4+1文献标志码: A文章编号:1002-1302(2020)13-0069-03
收稿日期:2019-08-12
基金项目:江苏沿海地区农业科学研究所基金(编号:YHS201610)。
作者简介:刘艳艳(1990—),女,山东日照人,硕士,助理研究员,主要从事水稻育种及抗病性研究。E-mail:1052233980@qq.com。
通信作者:严国红,研究员,主要从事水稻遗传育种研究。E-mail:2376308693@qq.com。水稻(Oryza sativa)作为世界上重要的粮食作物之一,为全球超过1/2的人口提供粮食来源[1]。由稻瘟病病菌(Magnaporthe grisea)引起的稻瘟病是水稻生产中最具有毁灭性的病害,在我国几乎所有水稻栽培地区都有稻瘟病害的发生,尤其是在南方稻区,曾多次出现稻瘟病大面积暴发事件,对我国的粮食生产造成了严重影响[2-3]。病原菌的生理小种多、变异强,能够在短时间内适应水稻抗病品种(系)的遗传抗性,导致很多抗病水稻品种(系)无法持久抗病,推广种植数年后就导致抗性减弱或完全丧失。因此,利用抗病基因选育稻瘟病抗性品种是防治稻瘟病的有效手段[4]。在传统水稻育种中,主要通过杂交系选结合抗病表型鉴定最终选育含有目标抗性的品种,依赖于育种者的育种经验,往往容易造成误差,导致抗性基因丢失,选择效率低且周期长[5]。因此,利用分子生物学技术明确主栽品种和新选育品种的基因型,同时结合传统育种手段,是抗病品种科学应用的前提。
迄今,水稻所有染色体上至少有69个抗稻瘟病位点被报道,包含84个主效基因,其中30多个抗稻瘟病基因已经被克隆[6]。许多与抗稻瘟病基因连锁的分子标记被开发,Pi-ta是最早被克隆出来的抗稻瘟病基因之一,位于水稻第12号染色体靠近着丝点附近的区域,包含2个外显子和1个长度为 1 463 bp 的内含子,编码1个长度为928个氨基酸的细胞质膜受体蛋白,其分子标记也相继被开发并广泛应用于水稻抗稻瘟病分子育种中[7]。水稻抗稻瘟病基因Pi9来源于小粒野生稻,Qu等克隆了Pi9抗稻瘟病基因,并将该基因定位在水稻第6号染色体上,结果表明,接种多个稻瘟病病菌生理小种后表现出较高的抗性[8]。华丽霞等利用PCR技术及电泳检测技术成功研发了稻瘟病抗性基因Pi2、Pi9及Piz-t的分子标记[9]。由此可见,利用水稻抗稻瘟病主效抗性基因Pi9的分子标记,有助于改良水稻对稻瘟病的抗性[10]。
本研究利用Pi-ta和Pi9抗病基因功能性分子标记,对32份粳稻育种骨干亲本进行抗稻瘟病基因型检测,旨在明确骨干亲本的抗病基因型,为下一步选育新的抗稻瘟病品种组合提供依据。
1材料与方法
1.1试验地点与材料
供试材料于2017年春季播种于江苏沿海地区农业科学研究所南洋试验农场(120°14′E,33°25′N)。供试材料为32份粳稻育种骨干亲本,由江苏沿海地区农业科学研究所提供,详见表1,其中已经审定的品种为科腾稻2号、泗稻2333、云粳稻1号、盐稻9号、扬育粳2号、苏10-100、盐稻1333和徐稻3号,其余材料为稳定的水稻品系。
1.2稻瘟病抗性基因Pi-ta和Pi9的分子标记
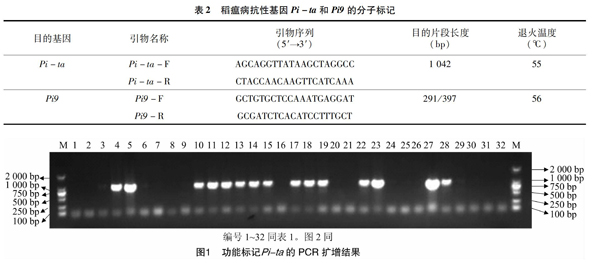
稻瘟病抗性基因Pi-ta、Pi9引物序列参照高利军等的研究结果[11],引物名称、序列及其目的扩增片段长度等信息见表2,引物由生工生物工程(上海)股份有限公司合成。
1.3稻瘟病抗性基因Pi-ta、Pi9的检测
采用十二烷基三甲基溴化铵(CTAB)方法提取水稻基因组DNA。以基因组DNA为模板进行基因片段扩增,PCR反应体系参照范方军等的方法[12]。反应产物经1%琼脂糖凝胶电泳分离后,用溴化乙锭染色,随后在紫外凝胶成像仪上观察并照相。
2结果与分析
2.1抗性基因Pi-ta的分子检测
用抗稻瘟病基因Pi-ta的特异性引物对32份供试粳稻材料进行PCR扩增。图1中的M为TaKaRa公司生产的DL2000 DNA marker,1~32对应表1中的品种(品系)。由图1可以看出,有15个水稻品种(品系)扩增出1条稍大于1 000 bp的特异性条带,分别是南粳5840、镇稻449、科腾稻2号、淮粳1309、泗稻2333、淮780、云粳稻1号、皖垦粳218、盐稻13180、扬育粳2号、苏10-100、宁3817、连粳15214、扬辐粳5208、浙粳20,表明所用水稻品种(品系)中含有抗稻瘟病基因Pi-ta。携带Pi-ta抗性基因的水稻品种(品系)约占32份水稻品种(品系)的46.88%,说明Pi-ta抗性基因已经在水稻骨干亲本中广泛存在。
2.2抗性基因Pi9的分子检测
用抗稻瘟病基因Pi9特异性引物对32份供试粳稻材料进行PCR扩增,图2中的M为TaKaRa公司生产的DL1000 DNA marker,1~32对应表1中的品种(品系)。由图2可以看出,只有4个水稻品种(品系)扩增出大小为291 bp的特异性条带,分别是江稻159、苏10-100、盐稻1333、徐稻3号。携带 Pi9 抗性基因的水稻品种(品系)占32份水稻品种(品系)的12.50%,说明Pi9抗性基因在水稻骨干亲本中的应用较少,应该加大对抗性基因Pi9的利用。
2.3抗性基因Pi-ta和Pi9的分布
由表3可以看出,在32份骨干亲本中,Pi-ta抗性基因的检出率为46.88%,Pi9抗性基因的检出率为12.50%,同时检测到Pi-ta和Pi9抗病基因的比例为3.12%,没有检测到Pi-ta和Pi9抗病基因的比例为43.75%。在骨干亲本中,Pi-ta单个基因的检出率较高,Pi9单个基因的检出率较低,同时含有Pi-ta和Pi9抗病基因的材料较少。因此,在以后的水稻抗病育种中,应该加强多个抗稻瘟病基因的聚合利用。
3讨论与结论
传统的水稻抗病育种主要依靠水稻品种在田间的发病程度进行筛选,这种育种方法耗时费力,而且容易受到环境的影响。稻瘟病病菌具有多个生理小种,且变异速度较快,一些抗病水稻品种推广 2~3年后表现出抗病性逐渐丧失的现象,因此,明确品种本身的抗性基因型对于品种的抗性改良尤为重要。随着抗稻瘟病基因的克隆及分子标记的开发,通过分子标记辅助选择检测水稻是否含有抗病基因,合理选择适宜在不同生态地区种植的抗性品种的方法,因具有准确、快速且不受环境影响等优点,已经被广泛应用于育种实践中。
近年来,国内外学者相继对水稻品种的抗瘟基因型开展了研究。张银霞等利用Pi-ta、Pi-b、Pi9对宁夏水稻种质资源进行了检测,为宁夏水稻分子标记辅助育种奠定了基础[13]。王军等研究发现,Pi-ta和Pi-b基因的联合效应与穗颈瘟抗性呈正相关,并且这种相关性比2个基因单独存在时与穗颈瘟抗性的相关性要紧密[14]。王生轩等研究发现,在河南地区,水稻稻瘟病抗病基因Pi9和Piz-t在育种中利用得较为广泛,Pi-ta利用得较少,今后河南省在水稻育种工作中应加强广谱稻瘟病抗性基因的聚合育种和综合利用[15]。马继琼等研究发现,贵州禾通过长期的人工和自然选择,Pi5和Pi-kh基因频率相对较高,而Pi9和Pi2频率较低[16]。范方军等研究发现,Pi-ta、Pi-b、Pi54是水稻育种中重要的抗稻瘟病基因,表明抗稻瘟病基因Pi-ta和Pi-b与穗颈瘟抗性存在显著相关性[12]。邹德堂等研究发现,在黑龙江地区水稻抗稻瘟病基因Pi9具有很高应用价值,含有Pi9基因的材料,苗期和分蘖期的平均发病级别均为2.4级,不含有Pi9基因的材料,苗期和分蘖期的平均发病级别分别为5.1级和5.4级[17]。在本研究中,Pi-ta检出率为46.88%,Pi9的检出率为12.50%,因此在今后的育种中,可以适当加强对Pi9基因的利用。
本研究中,骨干亲本广谱抗病基因Pi9在供试材料中分布得较少,抗性资源没有得到充分利用。因此,在以后的育种实践中,可以利用遗传多样性的方法对稻瘟病进行防治,将不同类型的抗稻瘟病品种种植在同一地区,这样可以避免抗稻瘟病品種在种植2~3年后其抗性逐渐丧失的现象,从而延长抗性品种的使用年限。此外,在后续研究中,应聚合不同抗性基因来提高抗性,避免单一抗病基因丧失。
参考文献:
[1]Liu J L,Wang X J,Mitchell T,et al. Recent progress and understanding of the molecular mechanisms of the rice-Magnaporthe oryzae interaction[J]. Molecular Plant Pathology,2010,11(3):419-427.
[2]刘万才,刘振东,黄冲,等. 近10年农作物主要病虫害发生危害情况的统计和分析[J]. 植物保护,2016,42(5):1-9,46.
[3]何秀英,刘新琼,王丽,等. 稻瘟病新隐性抗病基因pi55(t)的遗传及定位[J]. 中国科学(生命科学),2012,42(2):125-141.
[4]李旭升,向小娇,申聪聪,等. 水稻重测序核心种质资源的稻瘟病抗性鉴定与评价[J]. 作物学报,2017,43(6):795-810.
[5]王伟舵,于俊杰,聂亚锋,等. 2011—2014年江苏省稻瘟病菌种群动态及毒力变化[J]. 江苏农业学报,2015,31(2):285-289.
[6]蔡金洋. 分子标记在水稻抗稻瘟病育种中的研究进展[J]. 农业科技通讯,2018(6):4-5,9.
[7]王忠华,贾育林,吴殿星,等. 水稻抗稻瘟病基因Pi-ta的分子标记辅助选择[J]. 作物学报,2004,30(12):1259-1265.
[8]Qu S H,Liu G F,Zhou B,et al. The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of a multigene family in rice[J]. Genetics,2006,172(3):1901-1914.
[9]华丽霞,汪文娟,陈深,等. 抗稻瘟病Pi2/9/z-t基因特异性分子标记的开发[J]. 中国水稻科学,2015,29(3):305-310.
[10]周鹏,涂诗航,董瑞霞,等. 水稻抗稻瘟病基因分子检测及抗性评价[J]. 福建农业学报,2016,31(9):962-965.
[11]高利军,高汉亮,颜群,等. 4个抗稻瘟病基因分子标记的建立及在水稻亲本中的分布[C]//第1届中国杂交水稻水稻大会论文集,2010:294-298.
[12]范方军,王芳权,刘永峰,等. Pi-b、Pi-ta、Pikm和Pi54对水稻穗颈瘟的抗性评价[J]. 华北农学报,2014,29(3):221-226.
[13]张银霞,张敏,田蕾,等. 宁夏水稻品种抗稻瘟病基因 Pi-ta、Pi-b和Pi9的检测分析[J]. 江苏农业科学,2016,44(9):35-39.
[14]王军,杨杰,杨金欢,等. Pi-ta、Pi-b基因在江苏粳稻穗颈瘟抗性育种中的价值分析[J]. 华北农学报,2012,27(6):141-145.
[15]王生轩,李俊周,谢瑛,等. 河南粳稻抗稻瘟病基因Pi9、Pi-ta和Piz-t的分子检测[J]. 分子植物育种,2017,15(3):177-181.
[16]马继琼,孙一丁,杨奕,等. Pikh、Pi2、Pi9、Pi5 4个稻瘟病抗性基因在贵州禾中的分布[J]. 西南农业学报,2018,31(11):4-9.
[17]邹德堂,姜思达,赵宏伟,等. 广谱抗性基因Pi9在黑龙江省水稻品种中的分布[J]. 东北农业大学学报,2016,47(7):1-8.梁巩,张强,郭旭川,等. 不同来源血清对体外培养驴巨噬细胞的影响[J]. 江苏农业科学,2020,48(13):72-75.doi:10.15889/j.issn.1002-1302.2020.13.014
- 应用石墨烯的薄膜晶体管专利技术综述
- 浅谈超星读秀学术在肥料领域专利检索中的应用
- 国内农用无人机产业专利状况分析探讨
- 人乳头瘤病毒疫苗的发明专利分析
- 国产动画抄袭行为引发的版权侵权问题研究
- 论短视频的版权规制
- 网络音乐强制许可制度
- 论企业商标动态实施中的保护与限制
- 公共领域视野下注册商标转让的制度重构
- 从具体案例中探讨主题名称的选取对权利要求保护范围的影响
- 移动机器人路径规划实现方法研究
- 基于Eclipse的交通本体查询系统应用研究
- 基于STM32的光伏并网逆变器设计
- 基于Android系统的天气预报查询系统设计
- 河南省国家级产业化基地增至30家
- 2020中关村论坛全球科技创新智库论坛在京举办
- 河南省新型研发机构备案数量突破百家
- “科技列车怀化行”成果丰硕
- 21世纪中心参加金砖国家“海洋与极地科学”专题领域工作组第三届会议
- 2020全球硬科技创新大会在西安举办
- 终端污水处理系统节能减排研究
- 一起10 kV母线失压事件的原因分析及处理方案研究
- 地形图等高线采集重难点及特殊地貌表示方法
- 生态修复技术在秦安县城区段葫芦河生态治理中的应用
- 2018年4月濮阳春季首场暴雨天气过程的诊断分析
- superattainableness
- superattainablenesses
- superattainably
- superattendant
- superattendants
- superattraction
- superattractions
- superattractive
- superattractively
- superattractiveness
- superattractivenesses
- superaudible
- superauditor
- superauditors
- superaverage
- superaverageness
- superaveragenesses
- superaward
- superawards
- superb
- superbad
- superbank
- superbeing
- superbeings
- superbelief
- r2014097200000854
- r2014097200000856
- r2014097200000857
- r2014097200000859
- r2014097200000861
- r2014097200000862
- r2014097200000863
- r2014097200000865
- r2014097200000867
- r2014097200000868
- r2014097200000870
- r2014097200000871
- r2014097200000872
- r2014097200000873
- r2014097200000874
- r2014097200000876
- r2014097200000877
- r2014097200000878
- r2014097200000880
- r2014097200000882
- r2014097200000884
- r2014097200000886
- r2014097200000887
- r2014097200000888
- r2014097200000889