李易芯 万群 徐文君 葛静 曾东强 余向阳


摘要:通过测定不同代谢抑制剂和细胞色素P450s抑制剂对上海青吸收噻虫嗪的影响,探讨噻虫嗪的吸收及代谢机制,为农药施用及提高药效提供理论依据。在10 mg/L噻虫嗪水溶液中加入不同浓度的不同种类代谢抑制剂和P450s抑制剂,水培上海青1 d后测定植株体内噻虫嗪含量。10、50 μmol/L碳酰氰间氯苯腙和2,4-二硝基苯酚均能极显著降低上海青体内噻虫嗪含量,推测上海青吸收噻虫嗪过程中存在须要消耗能量的主动吸收。而含有10 mg/L胡椒基丁醚、1- 氨基苯并三唑和马拉硫磷的处理组中噻虫嗪含量极显著升高,推测上海青体内含有能代谢噻虫嗪的P450多功能氧化酶。上海青吸收噻虫嗪过程需要能量的供应,进入体内的噻虫嗪会被细胞色素P450代谢。
关键词:上海青;噻虫嗪;代谢抑制剂;细胞色素P450抑制剂;主动吸收
中图分类号:TQ450.1?文献标志码: A
文章编号:1002-1302(2020)04-0172-04
收稿日期:2018-11-19
基金项目:国家自然科学基金(编号:31601660,31772198)。
作者简介:李易芯(1993—),女,重庆人,硕士,从事农产品质量安全研究。E-mail:18883252739@163.com。
通信作者:曾东强,博士,教授,从事有害生物化学防治研究,E-mail:zengdq550@163.com;余向阳,博士,研究员,从事污染物残留代谢调控及其机理研究,E-mail:yuxy@jaas.ac.cn。
农药在防治作物病虫草害、保障农业增产增收方面发挥了重要作用。我国是农药使用大国,但农药利用率比较低,大量未发挥作用的农药直接进入到农田环境,对农业环境和农产品安全造成极大的威胁。作物通过根系将残留于土壤中的农药吸收,经过体内的迁移、转化后将农药分配在整个植物体内[1-2]。噻虫嗪是全球应用最为广泛的新烟碱类杀虫剂之一,常作为种子包衣剂用于多种农作物的害虫防治,其施用后可迅速被植物吸收,并传导到植株各部位。研究农作物对噻虫嗪的吸收和转运机制,可为噻虫嗪的科学合理使用和保障农产品安全提供科学依据。
在生物体中,各种物质进出细胞的过程须要通过蛋白组成的特殊途径介导才能完成。膜蛋白介导的分子或离子的跨膜运输过程可分为主动运输和被动运输两大类。主动运输是一种耗能过程,主要由线粒体提供能量。在真核细胞线粒体中,氧化磷酸化过程是物质在体内氧化时释放的能量通过呼吸链供给二磷酸腺苷(adenosine diphosphate,简称ADP)与无机磷合成腺苷三磷酸(adenosine triphosphate,简称ATP)的偶联反应。线粒体氧化磷酸化的电子传递链位于线粒体内膜上,内膜两侧具有电化学势梯度,使质子在ATP合成酶辅助下用于合成ATP,而ATP是细胞内流通的能量通货,其释放的能量可以供给各项生命活动。
2,4-二硝基苯酚(2,4-dinitrophenol,简称DNP)和碳酰氰间氯苯腙(carbonylcyanide-m-chlorophenylhydrazone,简称CCCP)是氧化磷酸化的解偶联剂,抑制ATP生成。DNP和CCCP增大了线粒体内膜的通透性,破坏质子梯度的形成,使电子传递链崩塌,由电子传递产生的能量以热被释出,并且它还会引起线粒体中ATP大量水解[3-5]。如在温度为20 ℃时,加入0.1 mmol/L CCCP,在20~30 s以内ATP浓度呈指数级下降,而膜电位在2~3 s 内呈指数级下降[6]。
细胞色素P450s酶系(cytochrome P450,简称CYPs)是一类广泛分布于动植物和微生物中的多功能氧化酶,含有單氧合酶的超家族,是一组最重要的代谢酶,即具有生物合成功能,还参与内源物质和外源物质的解毒代谢,如催化药物、杀虫剂、植物毒素等的氧化代谢,还能催化还原解毒反应[7-10]。胡椒基丁醚(PBO,增效醚)、1-氨基苯并三唑(1-aminobenzotriazole,简称ABT)和马拉硫磷是高效、无毒的广谱性P450s酶抑制剂,能够强烈抑制微粒体P450酶的活性,对细胞色素P450s酶家族具有非特异性的抑制作用[11-15]。PBO作为P450酶的抑制剂,已被广泛用作于有效的杀虫剂增效剂,通过对昆虫体内多功能氧化酶系的抑制作用,减少杀虫剂在昆虫体内的降解,从而达到增效作用[16]。马拉硫磷(malathion)是世界上使用最频繁的有机磷硫酸酯(OPT)杀虫剂之一,它通过羧酸酯酶的快速降解与细胞色素P450在毒性代谢物催化下形成竞争[17]。以噻虫嗪为材料,初步探讨上海青(Brassica chinensis L.)吸收农药是否存在主动吸收过程及其在植物体内的代谢机制。
1?材料与方法
1.1?仪器与试剂
试验主要的仪器与试剂有WH-3微型漩涡混合仪(上海沪西分析仪器厂有限公司)、Five go型实验室pH计[梅特勒-托利多仪器(上海)有限公司]、JJ223BC型电子天平(常熟市双杰测试仪器厂)、Agilent Technologies 1260-6410液相色谱串联质谱联用仪[安捷伦科技(中国)有限公司];噻虫嗪标准品、乙腈、霍格兰(Hoagland)全营养液。
噻虫嗪原药由济南绿霸农药有限公司提供;上海青由南京绿领种业有限公司提供。
1-2,4-二硝基苯酚(DNP)[酷尔化学科技(北京)有限公司]、碳酰氰间氯苯腙(CCCP)(合肥博美生物科技有限责任公司)、胡椒基丁醚(PBO)(上海麦恪林生化科技有限公司)、氨基苯并三唑(ABT)(上海源叶生物科技有限公司)、马拉硫磷(上海阿拉丁生化科技股份有限公司)。
1.2?试验设计
取上海青种子移入装有蛭石基质土的穴盘中,每穴1粒种子,培养期间用霍格兰全营养液[pH值为5.8~6.0,电导率(EC)为1.2~1.3 mS/cm]浇灌,20 d左右长至3叶1心,选取长势一致的幼苗待用。种植期间,以LED白光为生长光源,光照度为(130±5) μmol/(m2·s),光暗周期为16 h/8 h,环境温度为25 ℃。
用去离子水将上海青幼苗冲洗干净,吸水纸吸干植株表面水分,放入含不同处理液的50 mL玻璃离心管中。水培用的营养液为霍格兰全营养液,每天定期向玻璃瓶里补充霍格兰全营养液,3 d后检测整株上海青中的噻虫嗪含量,每个处理重复5次。
1.3?上海青体内噻虫嗪含量的测定
1.3.1?提取与净化
将每株上海青洗净,用电子天平分别称鲜质量,剪碎,放入10 mL塑料离心管中,放入1颗小钢珠振荡5 min粉碎均匀样品。加入5 mL 乙腈涡旋均匀,超声提取10 min再振荡提取30 min,加入1.0 g NaCl,充分涡旋,5 000 r/min离心5 min。取上层提取液2 mL加入预先装有50 mgN- 丙基乙二胺(primary secondary amine,简称PSA)、30 mg 石墨化碳黑(GCB),150 mg无水MgSO4的离心管(10 mL)中,涡旋30 s,5 000 r/min离心5 min,取上清过0.22 μm有机滤膜,滤液经HPLC-MS检测。
1.3.2?色谱质谱条件
(1)液相色谱条件:AgilentSB-C18色谱柱(2.1 mm×150 mm,3.5 μm);柱温为25 ℃;流动相为乙腈/水=7/3(体积比);流速为0.2 mL/min;进样量为5 μL。(2)质谱条件:电喷雾离子源,正离子源(electrospray ionization,简称ESI),离子源温度为150 ℃。采用多反应离子监测模式(multiple reaction monitoring,简称MRM),以质荷比(m/z) 292.1,2个子离子m/z 211.2、181.1进行定性分析,以母离子m/z 292.1和响应值最高的子离子m/z 211.1进行定量分析。噻虫嗪的保留时间为2.45 min,采用外标法定量。
1.4?方法的线性相关性、准确度及精密度
1.4.1?方法相关性
用电子天平准确称量0.103 g噻虫嗪标准品,用色谱级乙腈溶解,定容至100 mL,配制成浓度为1 000 mg/L的噻虫嗪标准溶液,放置在4 ℃冰箱中保存。将1 000 mg/L标准溶液稀释成0.01、0.02、0.05、0.10、0.50、2.00 mg/L等6个浓度,按“1.3.2”节中LC-MS的条件进样检测。以噻虫嗪溶液浓度为横坐标,峰面积为纵坐标作图,得标准曲线为y=213.4124 38x-840.774 919,r2=0.999 633 29。由此可见,在0.01~2.00 mg/L浓度范围内,峰面积与噻虫嗪浓度具有良好的线性关系。
1.4.2?回收率与精密度
每株空白上海青样品用钢珠匀浆后,根据样品质量添加5、10、20 mg/kg 水平的噻虫嗪标准溶液,每个添加量5个重复,涡旋均匀后放置1 h。然后按“1.3.1”节加入乙腈进行提取、净化,检测并计算添加回收率,结果如表1所示。在上海青植株中平均添加回收率为86.9%~93.0%,相对标准差为0.61%~1.20%。可见,此方法适用于上海青植株内噻虫嗪含量的检测。
1.5?数据分析
图中的每个数据点均是平均值。在95%的置信水平下,通过单因素方差法分析数据之间的显著差异。数据的表达方式为“平均值±标准差”;图像绘制采用Microsoft Excel 2016软件完成。
2?结果与分析
2.1?代谢抑制剂对噻虫嗪吸收的影响
由图1可见,上海青在含有10 μmol/L CCCP的噻虫嗪水溶液中培养1 d后,植株体内的噻虫嗪含量减少26%,50 μmol/L CCCP处理组减少24%;在含有1 μmol/L DNP的噻虫嗪水溶液中培养1 d后噻虫嗪含量减少21%,含10 μmol/L DNP处理组减少27%。于添加代谢抑制剂的溶液中水培的上海青体内噻虫嗪含量较对照均显著降低,可见CCCP、DNP的添加会导致上海青体内噻虫嗪含量降低。试验中上海青通过根系吸收噻虫嗪,说明其受到抑制是体内噻虫嗪含量降低的主要原因。CCCP和DNP是解偶联剂,推测其通过抑制ATP生成抑制根系吸收。
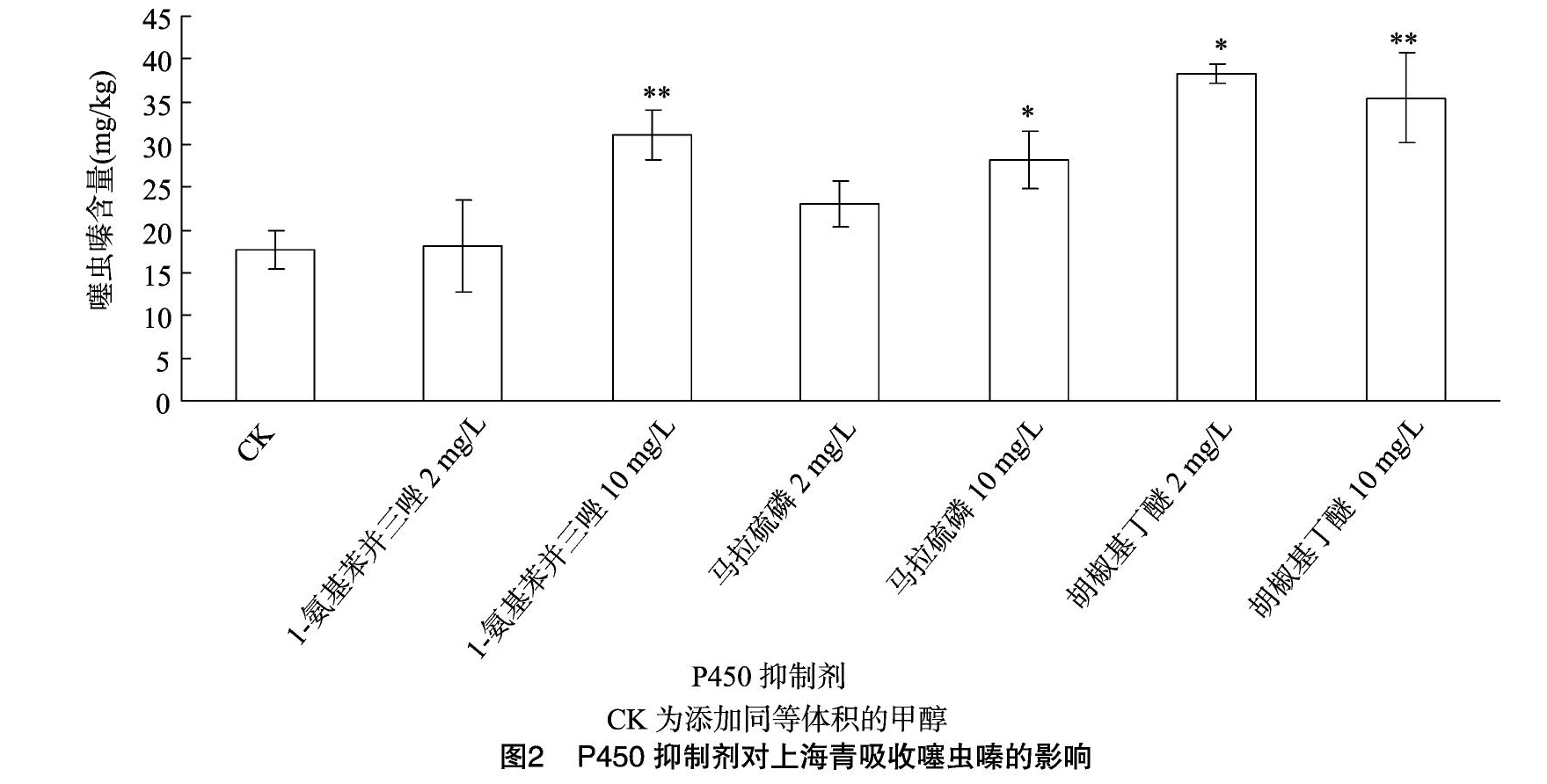
2.2?P450抑制剂对噻虫嗪吸收的影响
由图2可见,在含有10 mg/L 1-氨基苯并三唑(ABT)、马拉硫磷的噻虫嗪水溶液中培养1 d后,上海青体内噻虫嗪含量分别增加76%、59%;加入2、10 mg/L胡椒基丁醚(PBO)均能使上海青内的噻虫嗪含量加倍。ABT、马拉硫磷和PBO均是细胞色素P450的抑制剂,可见上海青体内存在的細胞色素P450且对外源添加物噻虫嗪在其体内的积累起着不可忽视的作用,在此过程中,PBO的效果最显著。
3?结论
Felle等用含有CCCP的溶液处理鹿角苔发现,当CCCP浓度高于0.3 μmol/L时,鹿角苔细胞中的ATP水平极显著地降低[6];解偶联剂CCCP使细胞内ATP合成受阻,抑制主动外排泵的能量来源,从而抑制药物外排[18-19]。杨根平等用呼吸解偶联剂2,4-二硝基苯酚(DNP)处理大豆下胚轴,同样发现ATP合成减少,且膜透性增大[20];林毅雄等用DNP处理采后的龙眼果实发现,DNP处理的龙眼果皮ATP含量极显著降低且下降速度变快,由于DNP处理使细胞内能量亏缺,诱导呼吸速率上升,也增加了细胞膜透性[21]。在本试验中,加入CCCP和DNP处理会使上海青体内噻虫嗪含量极显著降低。由此推测,CCCP、DNP抑制上海青体内的ATP水平,而吸收噻虫嗪的过程属于需要ATP提供能量的主动吸收过程,使上海青吸收噻虫嗪过程受阻。
1969年Frear等首次在棉花幼苗细胞中发现,能代谢灭草隆的多功能氧化酶,直接证明了植物细胞色素P450s酶系在除草剂中的解毒代谢作用[22]。Kawahigashi等将含有3种细胞色素P450s单加氧酶基因的质粒pIKBACH转入水稻,转基因水稻对莠去津、异草胺和诺氟沙星以及3种除草剂的混合物具有耐受性,且能降低水稻体内的除草剂残留[23]。
辛雪成喷施噻虫嗪的同时配合施用PBO防治棉蚜虫,噻虫嗪敏感品系增效比仅为0.95,而噻虫嗪抗性品系增效比提高3倍,PBO极显著地增加了噻虫嗪对棉蚜的毒性,证实P450解毒酶在抗噻虫嗪棉蚜品系的体内起到主要解毒代谢作用,是重要解毒酶,进一步说明棉蚜对噻虫嗪的解毒代谢作用与细胞色素P450酶相关[14]。胡椒基丁醚(PBO)和1-氨基苯并三唑(ABT)可明显抑制氯丹的降解,且抑制作用随抑制剂浓度的升高而增强[24]。ABT能够抑制西玛津在黑麦草(Lolium rigidum)体内的代谢[25]以及绿麦隆和异丙隆在小麦植株体内代谢[26]。Elmore等发现,马拉硫磷比ABT更能增强苯唑草酮对匍匐翦股颖的毒性[27]。然而,这3个P450酶抑制剂对植物吸收外源农药的影响仍未明确。本试验结果显示,3种P450抑制剂均能促进上海青对噻虫嗪的吸收,推测其是通过抑制上海青体内P450酶代谢噻虫嗪的活性来增加噻虫嗪在上海青体内的含量。
参考文献:
[1]卜元卿,孔?源,智?勇,等. 化学农药对环境的污染及其防控对策建议[J]. 中国农业科技导报,2014,16(2):19-25.
[2]Bell R M,Sferra P. Higher plant accumulation of organic pollutants from soils[R]. Cincinnati: US Environmental Protection Agency Risk Reduction Engineering Laboratory,1992.
[3]王吉秀,太光聪,祖艳群,等. 小花南芥根(Arabis alpinal var.parviflora Franch)对铅锌吸收的药理学研究[J]. 中国农学通报,2011,27(9):206-211.
[4]高?瑛,刘亚军,蔡欣然,等. 羰基氰氯苯腙调节非经典自噬的机制研究[J]. 中国生化药物杂志,2016,4(4):34-36,40.
[5]Dopson M,Lindstrm E B,Hallberg K B. ATP generation during reduced inorganic sulfur compound oxidation by Acidithiobacillus caldus is exclusively due to electron transport phosphorylation[J]. Extremophiles(Life Under Extreme Conditions),2002,6(2):123-129.
[6]Felle H,Bentrup F W. A study of the primary effect of the uncoupler carbonyl cyanide m-chlorophenylhydrazone on membrane potential and conductance in Riccia fluitans[J]. Biochimica et Biophysica acta,1977,464(1):179-187.
[7]趙?春,李银生,蒋美琳,等. 细胞色素P450与外源物的相互作用研究进展[J]. 中国现代应用药学,2014,31(8):1020-1024.
[8]汪思远,蒋世翠,王康宇,等. 植物细胞色素P450的研究进展[J]. 吉林蔬菜,2014(4):41-45.
[9]范银君,史雪岩,高希武. 新烟碱类杀虫剂吡虫啉和噻虫嗪的代谢研究进展[J]. 农药学学报,2012,14(6):587-596.
[10]Chen W,Lee M K,Jefcoate C,et al. Fungal cytochrome p450 monooxygenases:their distribution,structure,functions,family expansion,and evolutionaryorigin[J]. Genome Biology and Evolution,2014,6(7):1620-1634.
[11]谭利蓉. 阿特拉津胁迫下水稻细胞色素P450酶系的研究[D]. 南京:南京农业大学,2015.
[12]孙静娜,董?威,赵?帅,等. CCCP泵抑制剂对多重耐药及泛耐药鲍曼不动杆菌作用的研究[J]. 中国卫生检验杂志,2014(21):3159-3161.
[13]李久辉,程家高,邵旭升,等. 广谱增效剂PBO与P450酶作用模式的计算模拟研究[C]//中国化工学会农药专业委员会第十七届年会论文集,2016:399-403.
[14]辛雪成. 噻虫嗪抗性棉蚜筛选,交互抗性谱测定及差异转录组分析[D]. 长春:吉林大学,2016.
[15]王?佳. 过氧化氢对铜绿微囊藻的生态毒性:生长阶段效应与P450酶介导作用[D]. 杭州:浙江大学,2018.
[16]魏绪强,李秋红,周小洁. 增效醚在德国小蠊不同发育阶段对残杀威和毒死蜱增效作用的研究[J]. 中华卫生杀虫药械,2018(1):15-17.
[17]Buratti F M,Daniello A,Volpe M T,et al. Malathion bioactivation in the human liver:the contribution of different cytochrome p450 isoforms[J]. Drug Metabolism and Disposition(the Biological Fate of Chemicals),2005,33(3):295-302.
[18]Valdezate S,Vindel A,Echeita A,et al. Topoisomerase Ⅱ and Ⅳ quinolone resistance-determining regions in Stenotrophomonas maltophilia clinical isolates with different levels of quinolone susceptibility[J]. Antimicrobial Agents and Chemotherapy,2002,46(3):665-671.
[19]周明明,彭定輝,周铁丽,等. 主动外排在铜绿假单胞菌耐药中的作用研究[J]. 中华医院感染学杂志,2009,19(14):1786-1789.
[20]杨根平,荆家海. 渗透胁迫和呼吸抑制剂对细胞透性的影响[J]. 华北农学报,1993,8(2):50-53.
[21]林毅雄,林艺芬,陈?莲,等. 解偶联剂DNP处理对采后龙眼果实呼吸作用和细胞膜透性的影响[J]. 中国食品学报,2018(2):191-196.
[22]Frear D S,Swanson H R,Tanaka F S. N-demethylation of substituted 3-(phenyl)-1-methylureas;isolation and characterization of a microsomal mixed function oxidase from cotton[J]. Phytochemistry,1969,8(11):2157-2169.
[23]Kawahigashi H,Hirose S,Ohkawa H,et al. Phytoremediation of the herbicides atrazine and metolachlor by transgenic rice plants expressing human CYP1A1,CYP2B6,and CYP2C19[J]. Journal of Agricultural and Food Chemistry,2006,54(8):2985-2991.
[24]肖鹏飞,近藤隆一郎. 细胞色素P450抑制剂对白腐菌Phlebia lindtneri降解氯丹的影响[J]. 农药学学报,2012,14(5):515-520.
[25]Burnet M M,Loveys B R,Holtum J M,et al. Increased detoxification is a mechanism of simazine resistance in Lolium rigidum[J]. Pesticide Biochemistry and Physiology,1993,46(3):207-218.
[26]Cabanne F,Huby D,Gaillardon P,et al. Effect of the cytochrome P450 inactivator 1-aminobenzotriazole on the metabolism of chlortoluron andisoproturon in wheat[J]. Pesticide Biochemistry & Physiology,1987,28(3):371-380.
[27]Elmore M T,Brosnan J T,Armel G R,et al. Cytochrome P450 inhibitors reduce creeping bentgrass (Agrostis stolonifera) tolerance to topramezone[J]. PLoS One,2015,10(7):e0130947.
- 中小企业互联网融资风险防范体系研究
- 浅析金融学中大数据思维的应用
- 论保险业税务风险的有效应对
- 新企业会计准则下企业并购的会计处理方法研究
- 互联网金融背景下商业银行服务“三农”的创新策略研究
- 河南加快保险服务业发展问题研究
- 网红经济下的实体店营销策略研究
- 基于优衣库浅谈服装品牌的营销策略
- 大连地区大学生手机购物消费行为特征研究
- 高校公共安全突发事件与应急管理机制构建与创新
- 互联网时代中小企业市场营销的策略规划
- 加油站便利店营销策略研究
- 企业营销渠道冲突与战略管理
- 苹果手机营销中的文化因素
- 试论知识经济时代的市场营销策略
- 基于新媒体的汽车营销模式探讨
- 试论电商环境营销渠道与传统营销渠道的冲突
- 企业市场营销活动的项目化管理研究
- 基于互联网时代下市场营销策略的创新分析
- 基于国际贸易环境的市场营销策略革新研究
- 客户参与定制对购买意愿的影响
- 浅析二手交易平台“闲鱼”的经营之利
- 电子商务网络营销平台的安全性分析
- 欠发达地区发展跨境电商的困境与路径研究
- 计算机网络的防御技术探析
- menacing
- menacingly
- mend
- mended
- menders
- mending
- mendings
- mends
- mend your ways
- mend²
- mend¹
- menial
- more all out
- more all round
- more all time
- more anglo saxon
- more awe inspiring
- more back to back
- more bad tempered
- more ball bearing
- more beat up
- more big ticket
- more big time
- more black belt
- more bloody minded
- 镂花的窗格
- 镂花镌叶
- 镂襜
- 镂象
- 镂身
- 镂金
- 镂金铺翠
- 镂金错彩
- 镂金错影
- 镂金错采
- 镂金雕玉
- 镂金雕琼
- 镂银
- 镂错
- 镂雕
- 镂饰
- 镂骨
- 镂骨刻肌
- 镂骨铭心
- 镂骨铭肌
- 镂鸡子
- 镂鼎铭彝
- 镃
- 镃基
- 镃錤