柴智 黄琼 冯进 崔莉 黄午阳 李莹

摘要:以鱼肉理化性质、蛋白二级结构相对含量、质构特性、微观结构等为指标,探讨不同冻藏温度(-20、-30、-40 ℃)对鳜鱼贮藏过程中鱼肉品质变化的影响。结果表明,在不同冻藏温度下,鳜鱼肌肉的pH值、Ca2+-ATPase活性、总巯基含量均呈下降趋势,而表面疏水性指数和挥发性盐基氮含量均呈上升趋势;同时冻藏温度越低,样品的硬度、弹性、咀嚼性下降幅度越小;-40 ℃冻藏的鳜鱼样品具有更高的α-螺旋含量,其蛋白变性程度较小,-20 ℃冻藏的样品蛋白质无序程度明显增加。扫描电镜结果表明,不同冻藏条件下鱼肌纤维均发生一定变形,-20 ℃冻藏条件下的肌纤维结构遭到的破坏最严重,而-40 ℃冻藏条件下的肌纤维结构保持的相对比较完整。综合来看,-40 ℃冻藏时鳜鱼鱼肉品质各指标明显优于其他冻藏温度。
关键词:鳜鱼;鱼肉;冻藏;温度;品质;理化性质;蛋白二级结构;质构特性;微观结构
中图分类号: S984文献标志码: A文章编号:1002-1302(2020)13-0228-08
收稿日期:2020-06-11
基金项目:江苏省农业科技自主创新资金[编号:CX(19)2006]。
作者简介:柴智(1986—),女,安徽淮南人,博士,助理研究员,主要从事营养与食品加工领域研究。E-mail:sophia_chai@163.com。
通信作者:李莹,博士,副研究员,主要从事功能食品领域研究。E-mail:hijoly@163.com。鳜鱼(Siniperca chuatsi)别称桂花鱼,属鲈形目鱼旨科鳜鱼属,其肉质细腻,肌间少刺,营养丰富,是我国名贵的淡水鱼类[1]。鳜鱼经济价值高,近年来已突破繁养技术难关被广泛养殖,但鳜鱼暂养耗氧量大不易存活,这导致了大量鳜鱼不能被市场及时消化、积压于市,一旦失去食用价值将造成极大的经济损失,严重时将污染环境破坏水质。随着人们生活水平的提高,对水产品品质的要求也逐步提升,因此亟需采取保鲜措施以减缓鳜鱼鱼肉品质的劣变。寻找合适的贮藏方式,不仅能从原料上解决鳜鱼浪费的问题,同时对于保证鳜鱼的新鲜口感和质量安全也具有重要意义。
水产品常用的保鲜方式有低温保鲜、气调保鲜和高压静电场保鲜等技术,其中低温保鲜是应用最广泛的保鲜技术。低温冻藏保鲜技术是采用低温的手段把水产品的中心温度降至-15 ℃以下,并且在冷冻条件下贮藏和流通的一种保鲜措施[2]。目前已有利用冻藏技术对鲢鱼[3]、秋刀鱼[4]、河豚鱼[5]等进行保鲜的研究。低温冻藏可抑制水产品内微生物生长和内源酶的分解,从而延长产品的货架期,保持鱼肉品质。国内外研究表明,在冻藏温度、冻藏时间、冻结速率等条件影响下,鱼肉会在冻藏过程中发生蛋白质变性在内的多重理化反应,其中冻藏温度是最为关键的因素。
本研究选取不同冻藏温度(-20、-30、-40 ℃),以冻藏期内鳜鱼肌肉的pH值、Ca2+-三磷酸腺苷酶(ATPase) 活性、总巯基含量、表面疏水性指数、挥发性盐基氮含量、总蛋白二级结构相对含量、质构特性、微观结构等为指标,研究不同冻藏温度对冻结鳜鱼品质的影响,以期为鳜鱼的低温贮藏和保鲜提供理论依据和技术支撑。1材料与方法
1.1试验材料
鲜活鳜鱼,购于江苏省南京市嘉鸿水产商行,质量为(500±50) g/尾。将鲜活鳜鱼置于含冰的泡沫箱内,并在30 min内运回实验室。试验于2018年4—5月在江苏省农业科学院农产品加工研究所实验室进行。
三羟甲基氨基甲烷(Tris)、5,5′-二硫代双-硝基苯甲酸(DTNB)、8-苯胺基-1-萘基磺酸盐(ANS)(Sigma公司);二辛可宁酸(BCA)法蛋白定量测试盒、超微量Ca2+-ATP酶测试盒(南京建成生物工程研究所);磷酸氢二钠、磷酸二氢钠、氯化钠、氯化钾、戊二醛、盐酸、尿素等(国产分析纯)。
1.2试验方法
1.2.1材料预处理、冻藏及解冻去除鳜鱼的内脏、鱼皮和鱼骨后,用流水洗净后沥水10 min,剖片、整形,将鱼肉切分成3 cm×3 cm×3 cm大小的鱼块备用。将切好的鱼块随机分成3组,分别于低温冰箱中速冻至中心温度达到-20、-30、-40 ℃,然后分别置于-20、-30、-40 ℃条件下冻存。冻藏过程中定期取适量冻鱼块置于4 ℃冰箱中解冻2 h,解冻后的样品用于各项指标的测定。
1.2.2pH值的测定取鳜鱼鱼肉5 g,加入9倍体积的去离子水,12 000 r/min匀浆30 s,10 000 g离心10 min后取上清液测定pH值。
1.2.3肌原纤维蛋白的提取及Ca2+-ATPase活性的测定肌原纤维蛋白的提取参照Yang等的方法[6],取10 g鱼肉加入2倍体积的去离子水,充分匀浆后离心20 min(10 000 r/min、4 ℃),弃上清液,沉淀加去离子水重复上述步骤提取1次。收集沉淀,加入20 mL冷却的0.05 mol/L磷酸缓冲液(pH值为7.2,含0.6 mol/L NaCl),充分勻浆后,4 ℃、10 000 r/min 离心20 min,保留上清液并重复提取1次,合并2次上清液。蛋白质含量采用BCA试剂盒测定。Ca2+-ATPase活性的测定参照超微量 Ca2+-ATP 酶活性试剂盒操作说明书进行。
1.2.4总巯基含量的测定参照Benjakul等的方法[7],取0.5 mL肌原纤维蛋白溶液(蛋白浓度为 4 mg/mL),加入4.5 mL Tris-HCl 缓冲液[0.2 mol/L,pH值为6.8,含8 mol/L尿素,10 mmol/L乙二胺四乙酸(EDTA)和2%十二烷基硫酸钠(SDS)]。取 1 mL 上述溶液与100 μL 0.1% DTNB混合,于 40 ℃ 保温25 min后,在412 nm处测定溶液吸光度。以0.6 mol/L KCl溶液(pH值为7.0)代替样品作为空白对照。按照以下公式计算总疏基含量:
总疏基含量(nmol/mg)=[(D×n)÷(ε×ρ)]×106。
式中:D为412 nm波长处的吸光度;n为稀释倍数;ε为摩尔吸光系数,为13 600 L/(mol·cm);ρ为蛋白质质量浓度(mg/mL)。
1.2.5表面疏水性测定采用ANS荧光探针法测定表面疏水性[8]。首先用0.1 mol/L、pH值为7.0的磷酸缓冲液配制8 mmol/L的ANS溶液。然后用 10 mmol/L pH值为6.0的磷酸缓冲液(含 0.6 mol/L NaCl)将肌原纤维蛋白溶液稀释至0.125、0.250、0.500、1.000 mg/mL。取4 mL各浓度的肌原纤维蛋白溶液与30 μL ANS溶液混合,测定ANS-蛋白结合体的荧光强度(激发波长为 374 nm、发射波长为485 nm),以荧光强度对肌原纤维蛋白溶液质量浓度作图,计算曲线斜率即为蛋白表面疏水性指数。
1.2.6挥发性盐基氮(TVB-N)含量的测定TVB-N 含量的测定采用GB 5009.228—2016《食品安全国家标准食品中挥发性盐基氮的测定》中半微量定氮法进行。
1.2.7傅里叶红外光谱的测定取3 g鱼肉,加入30 mL 10% SDS溶液充分匀浆后,85 ℃水浴加热 1 h,10 000 g离心15 min,收集上清液中的总蛋白。取3 mg总蛋白与200 mg溴化钾混合,烘干,研磨后均匀压片。傅里叶红外光谱测定条件:分辨率为4 cm-1,扫描次数为32次。利用Peakfit软件对图谱进行去卷积和曲线拟合分析,根据各子峰的积分面积及其指认关系计算蛋白质二级结构的相对百分含量。
1.2.8质构特性的测定采用质构仪测定鱼肉的硬度、弹性和咀嚼性。将鱼肉切成20 mm×25 mm×30 mm大小,质构仪条件:平底柱形探头P/30(直径为30 mm),探头下降速率为3 mm/s,测试速率为 1 mm/s,返回速率为1 mm/s,压缩程度为50%。
1.2.9扫描电镜的测定将鱼肉切成1 mm3左右,在2.5%的戊二醛溶液中固定2 h以上,用 0.1 mol/L 磷酸缓冲液漂洗3次,每次15 min。然后用30%、50%、70%、80%、90%的乙醇溶液依次脱水15 min,用无水乙醇漂洗2次,每次10 min。将脱水后的样品放入叔丁醇中浸泡2 h以上后冷冻干燥。最终样品喷金后通过扫描电镜进行观察。
1.3数据处理
所有试验均重复3次,试验结果以平均值±标准差来表示,使用SPSS 18.0软件进行方差分析,试验数值间以ANOVA法进行显著性检验,选取P<0.05为显著水平。
2结果与分析
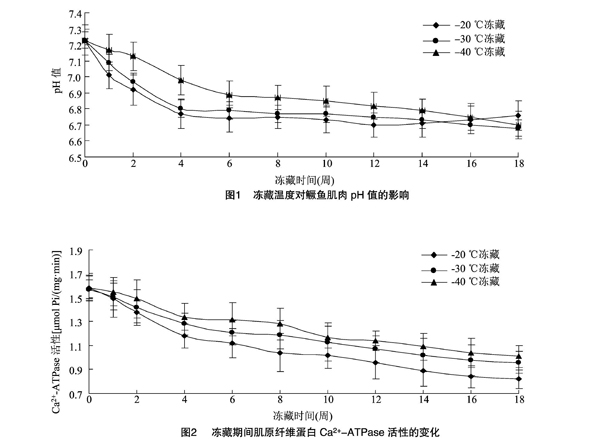
2.1冻藏温度对鳜鱼肌肉pH值的影响
由图1可知,在18周冻藏期内,冻藏鱼肉的pH值整体呈下降趋势。前2周内,-20、-30 ℃冻藏条件下鱼肉pH值快速下降,而-40 ℃冻条件下鱼肉pH值下降较缓慢。这表明冻藏温度會对肌肉组织pH值的变化产生较大影响,较低的冻藏温度能够有效抑制微生物的生长和鱼体内源酶对肌肉中蛋白质的分解作用,使得鱼肉中产生较少的胺类化合物及其他代谢产物,使得pH值变化较为缓慢[9]。
2.2冻藏温度对肌原纤维蛋白Ca2+-ATPase活性的影响
由图2可知,3种冻藏温度下的鳜鱼肌原纤维蛋白Ca2+-ATPase活性随冻藏时间的延长均呈下降趋势,尤其是-20、-30 ℃冻藏条件下,前2周内Ca2+-ATPase活性急速下降。18周冻藏期结束后,-20、-30、-40 ℃冻藏下的肌原纤维蛋白的 Ca2+-ATPase活性分别为0.82、0.96、1.01 μmol Pi/(mg·min),比新鲜鱼肉肌原纤维蛋白的 Ca2+-ATPase 活性分别降低48.1%、38.9%、36.1%。-20 ℃ 冻藏条件下的样品Ca2+-ATPase活性下降速率明显比较大,而-30、-40 ℃冻藏条件下的样品Ca2+-ATPase活性下降速率相对较小,且冻藏8周后变化趋于平稳。
有研究得出了相似的结论。Xiong 等研究发现,在-18 ℃冻藏30 d后,草鱼肉蛋白的 Ca2+-ATPase 活性随冻藏时间的延长最终降低了71.4%[10]。在冻藏过程中,肌球蛋白球状头部构象发生改变或者相互聚集,是造成肌原纤维蛋白 Ca2+-ATPase 活性下降的主要原因。冻藏温度对肌原纤维蛋白Ca2+-ATPase活性的下降速率有很大影响,冷冻贮藏温度越高,Ca2+-ATPase活性下降速率越大。
2.3冻藏温度对总巯基含量的影响
由图3可知,冻藏初始时蛋白质总巯基含量为124.2 nmol/mg,不同贮藏温度下鳜鱼样品的总巯基含量均随冻藏时间的延长而逐渐下降。在18周冻藏期结束后,-20、-30、-40 ℃冻藏温度下样品的总巯基残留量分别为冻藏初始值的56.5%、68.3%、74.8%。试验结果表明,在较低的冻藏温度下产生的冰晶相对较小,小冰晶对蛋白质空间结构的机械破坏力也较小,因而可以更好地保护蛋白质中的巯基不被氧化。
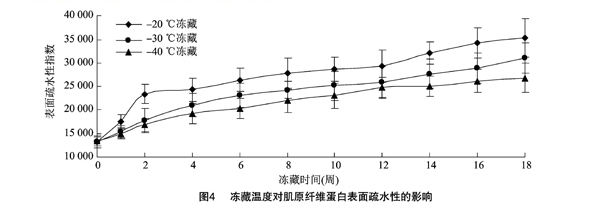
2.4贮藏温度对表面疏水性的影响
由图4可以看出,3种冻藏温度下肌原纤维蛋白的表面疏水性均随冻藏时间的延长而明显增强,且-20 ℃冻藏温度下样品表面疏水性上升趋势比较明显。冻藏12周后,不同冻藏温度下样品间表面疏水性差异明显,18周后蛋白质表面疏水性指数分别是初始值的2.69、2.36、2.00倍。
2.5冻藏温度对挥发性盐基氮含量的影响
不同冻藏温度下鳜鱼鱼肉贮藏过程中TVB-N含量的变化如图5所示,3种冻藏温度下样品的TVB-N含量均呈现上升趋势。在初始的4周内,鱼肉TVB-N含量上升较为缓慢;在冻藏中后期(8~16周),-20 ℃冻藏条件下样品的TVB-N含量上升速率明显增大,明显高于-30、-40 ℃冻藏条件下的样品。冻藏结束后,-20 ℃冻藏样品的TVB-N含量达到24.98 mg/100 g,而-30、-40 ℃冻藏样品的TVB-N含量上升速率较小,在冻藏结束后其TVB-N含量分别为16.20、11.53 mg/100 g。
2.6冻藏温度对蛋白质二级结构的影响
傅里叶红外光谱在分析蛋白质二级结构过程中的应用较为广泛,其吸收最强的区域为酰胺Ⅰ带(波数范围为1 600~1 700 cm-1),这个区域的变化主要由蛋白质分子多肽骨架CO的伸缩振动引起。二级结构中α-螺旋具有高度的稳定性,因此α-螺旋结构的相对含量可以作为蛋白质结构稳定性的评判标准。β-转角和无规卷曲则是蛋白质分子的无序结构。
不同冻藏温度下鳜鱼样品中蛋白质各二级结构的含量如表1所示。18周冻藏期结束后,-40 ℃贮藏的鳜鱼样品中α-螺旋含量为17.55%,β-折叠含量为40.00%,无规卷曲含量为17.62%,β-转角含量为24.83%。-40 ℃贮藏的鳜鱼样品具有更高的α-螺旋含量表明其变性程度较小。-20℃
冻藏的鳜鱼样品α-螺旋含量低于-30、-40 ℃冻藏组,而β-转角和无规卷曲含量则高于-30、-40 ℃ 冻藏组,说明-20 ℃冻藏时,鳜鱼的蛋白质发生了更严重的变性,α-螺旋结构逐渐解旋,部分转化为β-转角和无规卷曲,这与任丽娜等的研究结论[3,11]基本一致。而冻藏期结束后,-20 ℃条件下β-折叠含量降低,无规卷曲含量上升,说明鳜鱼肌肉的β-折叠结构进一步被破坏,蛋白质的无序程度增加。
2.7冻藏温度对质构特性的影响
硬度、弹性、咀嚼性等质构特性指标通常用来评价肌肉的品质[12]。硬度表示肉品达到一定程度形变所需要的力,也可以反映出人的触觉;弹性指外力作用于肉品时致其形变,去除外力后恢复形变的程度;咀嚼性指肉品咀嚼致可吞咽时所做的功[13]。冻藏过程中肌肉蛋白的变性改变了蛋白质的空间结构,使蛋白和水分子间的作用力减弱,原本存在于肌细胞间隙中的水分流失,从而将影响肌肉的组织形态、质构特性和感官品质。
不同冻藏温度的鳜鱼样品在贮藏过程中,鱼肉的硬度随冻藏时间的延长而逐渐下降(图6-a),其中-20 ℃冻藏样品的硬度下降最快,由上述 Ca2+-ATPase 活性、蛋白质二级结构等试验结果可知,-20 ℃冻藏样品中肌球蛋白变性程度最大,硬度也随之下降最快,而更低温度下冻藏可以较好地保护蛋白质的空间结构,使鱼肉的硬度下降缓慢。随冻藏时间的延长,3种冻藏温度下的鱼肉弹性均呈现下降的趋势,其中-40 ℃冻藏温度下的样品弹性下降速度最为缓慢(图6-b)。鱼肉咀嚼性的变化趋势与弹性和硬度的变化趋势一致(图6-c)。总的来说,冻藏过程中肌肉蛋白质的结构变化造成了鳜鱼鱼肉质构的松散,而低温贮藏可以明显减缓硬度、弹性和咀嚼性的下降速度。
2.8贮藏温度对组织微观结构的影响
由图7可以看出,新鲜鳜鱼鱼肉样品在扫描电镜下的横切切面显示肌纤维结构完整,纹理清晰可见,肌肉组织排列整齐,彼此间结合紧密。不同冻藏温度处理的鳜鱼样品在冻藏结束后,肌纤维均有不同程度的形变。-20 ℃冻藏条件下,鱼肉横切肌纤维结构明显疏松,肌束细胞遭到严重破坏,纹理模糊。这可能是由于较高的冻藏温度会在细胞内形成较大、较粗糙的冰晶,这种较大的冰晶会导致肌细胞结构被破坏。-30、-40 ℃冻藏条件下,横切的鳜鱼鱼肉组织变化相对不大,肌纤维排列仍较整齐且紧密, 未出现严重的撕裂情况。从纵切组织
可以看出, 与新鲜鱼片相比, 3种冻藏温度下,鳜鱼鱼肉纤维之间均有空隙,组织结构疏松,-40 ℃冻藏的样品肌纤维结构相对较完整。
3结论与讨论
本研究以冻藏期内鳜鱼肌肉pH值、Ca2+-ATPase活性、总巯基含量、表面疏水性指数、TVB-N含量、总蛋白二级结构相对含量、质构特性、微观结构等为指标,研究了不同冻藏温度(-20、-30、-40 ℃)对鳜鱼鱼肉品质变化的影响,结果表明,随着冻藏时间的延长,在不同冻藏温度下,鳜鱼肌肉的pH值、Ca2+-ATPase活性、總巯基含量均呈下降趋势,而表面疏水性指数和TVB-N含量均呈上升趋势。鱼肉pH值变化的快慢对冻藏鱼肉的品质会产生重要的影响,彭欢欢等研究发现-40 ℃低温冻藏下鱼肉pH值变化缓慢,有利于品质的保证[14],本研究结果与之类似。Ca2+-ATPase活性是反映肌球蛋白头部结构变化较为灵敏的指标[15]。冻藏温度越低,Ca2+-ATPase活性下降速率越小。巯基是肌原纤维蛋白中活性功能基团的重要组成部分[16]。本研究中总巯基含量的变化趋势与Benjakul等的研究结果[7]一致。表面疏水性增强的可能原因是冻藏促进鱼肉肌原纤维蛋白伸展,使更多的疏水基团外露[17]。TVB-N含量是反映水产品腐败程度的重要指标,较低的冻藏温度能更好地抑制微生物生长,从而减缓鱼肉的腐败,因此冻藏温
度越低,其TVB-N含量增幅越小。冻藏后各组样品的质构特性均呈下降趋势,冻藏温度越低,样品的硬度、弹性、 咀嚼性下降幅度越小。类似地,张南海研究发现不同冻结温度下,鲫鱼肉的硬度、弹性和咀嚼性均下降[18]。-40 ℃ 冻藏的鳜鱼鱼肉蛋白具有更高的α-螺旋含量,表明其蛋白变性程度较小;扫描电镜结果表明,3组冻藏条件下的鱼肉肌纤维均发生一定程度的变形,-20 ℃冻藏的样品肌纤维结构被破坏的最严重,-40 ℃冻藏的样品肌纤维结构保持的相对比较完整[18],这与王凤玉等的研究结果[4]是一致的。综合来看,-40 ℃ 冻藏温度下的鳜鱼鱼肉样品各品质指标变化趋势均明显优于-20 ℃冻藏组样品,冻藏温度越低,品质保持效果越明显。
参考文献:
[1]黄琼,方旭波,李莹,等. 鳜鱼下脚料酶解物对冷冻鳜鱼的保水作用[J]. 中国食品学报,2020,20(1):222-229.
[2]王子萌,苑宁,王琳琳,等. 保鲜技术在水产品中的应用研究[J]. 农产品加工,2017(4):46-48.
[3]任丽娜. 白鲢鱼肉肌原纤维蛋白冷冻变性的研究[D]. 无锡:江南大学,2014:6-7.
[4]王凤玉,曹荣,赵玲,等. 秋刀鱼-20 ℃、-30 ℃和-50 ℃冻藏过程中品质变化[J]. 食品研究与开发,2015,36(21):7-11.
[5]马妍,谢晶,周然,等. 冻藏温度对河豚鱼鱼肉微观结构及生化指标的影响[J]. 西北农林科技大学学报(自然科学版),2014,42(5):141-146.
[6]Yang F,Rustad T,Xu Y,et al. Endogenous proteolytic enzymes-a study of their impact on cod (Gadus morhua) muscle proteins and textural properties in a fermented product[J]. Food Chemistry,2015,172:551-558.
[7]Benjakul S,Sutthipan N. Muscle changes in hard and soft shell crabs during frozen storage[J]. LWT-Food Science and Technology,2009,42(3):723-729.
[8]Anacleto P,Teixeira B,Marques P,et al. Shelf-life of cooked edible crab (Cancer pagurus) stored under refrigerated conditions[J]. LWT-Food Science and Technology,2011,44(6):1376-1382.
[9]Lakshmanan P T,Antony P D,Gopakumar K. Nucleotide degradation and quality changes in mullet (Liza corsula) and pearlspot (Etroplus suratensis) in ice and at ambient temperatures[J]. Food Control,1996,7(6):277-283.
[10]Xiong G,Wei C,Ye L,et al. Effects of konjac glucomannan on physicochemical properties of myofibrillar protein and surimi gels from grass carp (Ctenopharyngodon idella)[J]. Food Chemistry,2009,116(2):413-418.
[11]朱孔辉.? 鳊鱼鱼糜在不同温度下保藏过程中理化特性变化及分析[D]. 上海:上海海洋大学,2009.
[12]Li X,Li J,Zhu J,et al. Postmortem changes in yellow grouper (Epinephelus awoara) fillets stored under vacuum packaging at 0 ℃[J]. Food Chemistry,2011,126(3):896-901.
[13]董開成. 不同低温预处理对小黄鱼贮藏过程中品质的影响[D]. 杭州:浙江大学,2015:53-54.
[14]彭欢欢,刘小莉,张金振,等. 不同冷冻方式对斑点叉尾鱼片品质的影响[J]. 食品研究与开发,2017,38(8):177-182.
[15]Benjakul S,Seymour T A,Morrissey M T,et al. Physicochemical changes in pacific whiting muscle proteins during iced storage[J]. Journal of Food Science,1997,62(4):729-733.
[16]潘锦锋,沈慧星,尤娟,等. 草鱼肌原纤维蛋白加热过程中理化特性的变化[J]. 中国农业大学学报,2009,14(6):17-22.
[17]Riebroy S,Benjakul S,Visessanguan W,et al. Acid-induced gelation of natural actomyosin from Atlantic cod (Gadus morhua) and burbot (Lota lota)[J]. Food Hydrocolloids,2009,23(1):26-39.
[18]张南海. 不同冻结方式、贮藏温度和解冻方式对彭泽鲫品质的影响[D]. 南昌:南昌大学,2018:31-32.李亚萍. 基于低酸法优化核桃油脱胶工艺参数[J]. 江苏农业科学,2020,48(13):236-239.doi:10.15889/j.issn.1002-1302.2020.13.048
- 对财务共享的认识
- 财务共享中心下的共享财务管理分析
- 新形势下企业财务会计与管理会计融合思考
- 大数据环境下企业财务的信息化建设
- 浅谈公立医院全成本核算体系的优化
- ERP财务软件在企业中的应用研究
- 内控制度的防火墙作用浅析
- 基于移动办公需求趋势下对企业应用智能财务管理的思考
- 大数据时代企业管理会计面临的挑战与解决对策
- 医院内部控制现状及改进措施
- 风劲扬帆正当时
- 金融供给侧结构性改革的内容与主攻方向
- 外币跨境资金池政策比较研究
- “互联网+”时代我国农业经济发展方式转变分析
- 论“互联网+”环境下如何做好水利经济发展规划
- 网络经济对我国国民经济发展的影响研究
- 绩效考核机制在医院经济管理中的核心作用
- 基于知识经济下的企业经济管理分析
- 建筑经济成本管理的问题分析
- 经济转型视域下企业的适度负债经营措施分析
- 关于加快县域民营经济高质量发展的几点思考
- 浅析我国房地产经济发展现状及未来发展趋势
- 县域经济和财政收入稳定增长研究分析
- 医院经济管理流程标准化分析
- 经济新常态下企业内控存在的问题与对策研究
- tingle
- tingled
- tingler
- tinglers
- tingles
- tingling
- tinglingly
- tinglish
- tinier
- tinies
- tiniest
- tininesses
- tinker
- tinkerdom
- tinkered
- tinkerer
- tinkerers
- tinkering
- tinkers
- tinkle
- tinkled
- tinkles
- tinkling
- tinlike
- tinned
- wǎ
- wǎn
- wǎng
- wǎnɡ
- wǒ
- wǔ
- wɑ
- wɑi
- wɑn
- wɑng
- wɑnɡ
- w世代
- w世代
- w 说话
- :-x
- x
- x-(
- [xchar[r201405004_fzhtk_f8cb]]
- [xchar[r201405004_fzktk_f8ac]][xchar[r201405004_fzshk_f8ad]]
- [xchar[r201405004_fzktk_f8e4]]
- [xchar[r201405004_fzktk_f9a9]]
- xi
- xia
- xian
- xiang