岳高峰 韩志强 薛志伟 刘辉 马丽婷

摘要:选取紫斑牡丹为试验材种,在0(CK)、30%、50%和70%不同遮阴度处理下,对牡丹的花期和叶片光合特性进行试验和研究分析。结果表明,不同的遮阴度下,光照对牡丹的花期时间和开花数量有明显的影响,对照处理的花期最早在30%遮阴度处理下,开花数量最多且质量最优。在CK和30%遮阴度处理下,牡丹光合日变化曲线为“双峰”型,有明显“光合午休”现象。随着遮阴度的增加,牡丹的净光合速率先升高后降低,蒸腾速率、气孔导度和胞间CO2浓度均呈现递减趋势。在固定的光合有效辐射梯度值下,30%遮阴度的净光合速率明显高于其他遮阴处理。牡丹叶片的光补偿点(Ic)和光饱和点(Isat)随着遮阴程度的增加均有明显下降。不同遮阴处理下光响应模型拟合结果中,非直角双曲线对斜率(α)拟合程度最优,指数方程模型对暗呼吸速率(Rd)和光补偿点(Ic)拟合程度最优,双曲线修正模型对最大净光合速率(Pmax)和光饱合点(Isat)拟合程度最优。
关键词:牡丹;遮阴;花期;光合特性;影响
中图分类号: S685.110.1 ?文献标志码: A ?文章编号:1002-1302(2020)17-0166-05
牡丹(Paeonia suffruticosa)属于毛茛科芍药属木本植物,性宜寒畏热,喜燥恶湿,为多年生落叶小灌木,在我国南北方广为种植。牡丹在每年的3—5月份开花,花期通常维持7~10 d。牡丹因其花色五彩斑斓、花型娇艳多姿和花香芬芳沁人心脾而广受群众喜爱,成为了我国优质的观赏花卉资源,具有极高的观赏价值[1]。除此之外,牡丹的食用和药用功能同样存在着巨大的经济价值和景观生态价值,因此它具有深远的市场开发潜力[2]。现代盛行的大田和园林式牡丹种植栽培技术极大地推动了牡丹观光产业的发展,并拉动和促进了当地经济发展。河南洛阳和山东荷泽每年在春季举行以牡丹观光为主题的旅游花会,吸引全球游客到此观光旅游,实现了旅游产业高速发展。
甘肃省临夏回族自治州(以下简称临夏州)地处青藏高原和黃土高原的过渡地带[3],地理海拔和气候均适宜种植紫斑牡丹。近年来,临夏州加快推进牡丹观光旅游产业步伐,在临夏市推出30里牡丹旅游观光长廊,并在每年春季举行“河州牡丹文化旅游节”,吸引省内外的游客来观光旅游,促进了当地旅游产业的快速发展,并取得了可观的经济社会效益。由于牡丹花期较短,时段比较集中,且对光照、气温等外部的环境因素比较敏感,因此研究牡丹生长发育所需的光照时长和光照度,延长牡丹盛花期时间,提升花期品质成为了牡丹旅游产业中重要的课题。
植物主要通过光合作用来存储物质能量[4]。光照对植物生长发育起着决定性的重要作用,也是最重要的外部环境影响因素之一[5]。光照主要通过光照度、光质和光照时间3种形式对植物的生长发育和形态变化进行调节和影响[6-7]。以园林、大田方式栽种的现代观光牡丹的花期及品质与光照条件密不可分。光照在植物的生长发育中起着重要的调控作用,特别是对植物的成花诱导和花芽分化方面起着不可替代的重要作用[8-9]。不同光照条件对牡丹的光合生理和形态响应又有着显著的影响[10]。本研究选取紫斑牡丹为试验材种,通过设置不同的遮阴度处理进行试验,在不同遮阴度处理下对牡丹叶片光合生理特性和牡丹花期进行分析,以期为园林、大田栽种牡丹的花期延长、品质提升提供科学支撑和数据参考。
1 材料和方法
1.1 试验地点
试验地点位于甘肃省临夏州振华牡丹园艺科技有限责任公司下属的河州紫斑牡丹示范园。示范园区地理位置为35°39′13″N、103°13′22″E,海拔高度为 1 956.8 m,属温带大陆性气候,年平均日照时长为2 474 h,年平均气温为7.1 ℃,年平均相对湿度为62%,年平均降水量为588.6 mm。
1.2 试验材料
选取6年株龄健壮且长势均匀的紫斑牡丹(Paeonia rockii)作为试验材种,于2018年3—2018年7月间在河州紫斑牡丹示范园的大田中进行试验。培养土质为普通园土(pH值为7.1)。
1.3 试验设计
3月20日,观测到牡丹进入显蕾物候期,在 20—21日搭建4个遮阴棚。遮阴棚南北走向,南北长4.0 m,东西宽2.5 m,高2 m,遮阴棚间距为 3 m,且将南北向设置为敞开,便于通风。在遮阴棚上固定不同透光率的黑色遮阳网,使用 TES-1335数字式照度计测定透光率,调整并设计遮光率分别为0、30%、50%和70%,分别对应于CK、T1、T2和T3共4种遮阴处理。牡丹遮阴处理期间的培养和浇灌按照往年常规方法进行。对牡丹圆桃期至花期的各个物候期进行观测记录。遮阴30 d后,对牡丹光合响应参数和叶绿素含量进行测量。
1.4 测定方法
1.4.1 牡丹物候期测定 为观察不同遮阴处理对牡丹花期的影响,在牡丹圆桃期至盛开期,连续观测记录圆桃期、平桃期、初开期、普开期和盛开期共5个物候期的开始时间,在花期时连续观测记录出现花序数量。
1.4.2 牡丹叶片光合色素测定 使用乙醇丙酮浸提比色法对牡丹叶片光合色素进行测定。并参考赵世杰等提供的方法[11-12]对牡丹叶片中的叶绿素a、叶绿素b含量进行测量。
1.4.3 光合日变化响应曲线测定 4月18—24日,选取天气晴朗且静风无云的日子,使用美国 LI-COR公司的LI-6400XT便携式光合测量系统对光合参数进行测定。主要测定参数为净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和细胞间CO2浓度(Ci)。测定时,在每个处理中选取牡丹3株,每株中选取顶中下面第3张叶片,共取3张叶片,并进行3次重复。测定时间为08:00—18:00,每隔2 h测定1次,每次测定在30 min内完成。
1.4.4 光响应曲线的测定 4月19日08:00—11:00 对牡丹叶片光合指标进行测定。测定时,先进行充分光活化处理[时长为30 min,光照度为 2 000 μmol/(m2·s)],使气孔完全张开。使用开放气路,利用发光二极管(LED)红蓝光源,控制CO2浓度为400 μmol/(m2·s),并设置光照度分别为 2 000、1 700、1 500、1 200、1 000、800、600、400、200、100、50、20、0 μmol/(m2·s)共13组光照有效辐射梯度,测定Pn,制作Pn-光合辐射(PAR)响应曲线。测定中设置自动取值间隔为最小等待3 min,最大 5 min。在响应曲线拟合时,参照Miko等的直角双曲线模型[13]、Marshall等的非直角双曲线模型[14-15]、叶子飘等的双曲线修正模型[16]和Kupp等的指数方程模型[17]进行数据拟合,并挑选出最佳拟合模型。
1.5 数据处理与分析
使用Excel 2010对试验数据进行处理,使用SPSS 21进行数据分析,使用Sigma Plot 12绘制图形。
2 结果与分析
2.1 不同遮阴处理对牡丹花期的影响
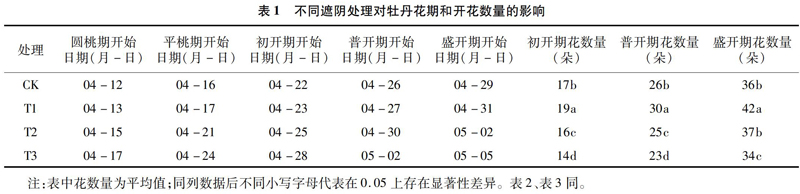
由表1可以看出,不同遮阴处理下,随着遮阴度的增加,牡丹物候期延迟。70%遮阴度处理的圆桃期比CK延迟了5 d,到盛花期时则延迟了 7 d,但30%遮阴度处理与CK的物候期开始时间差异较小。随着遮阴度的增加,牡丹开花数量呈现出先增后减趋势。在初开期、普开期和盛开期,30%遮阴度处理的开花数量都最高。
2.2 不同遮阴处理对牡丹叶片叶绿素含量的影响
通过分析不同遮阴处理下对牡丹叶绿素含量的影响(表2),可以看出,不同遮阴处理对叶绿素含量的影响存在着较大差异。随着遮阴度的增加,叶绿素a、叶绿素b、总叶绿素含量均呈现升高的趋势,而叶绿素a/b的变化规律则相反,这与周曙光等的研究结果[18-20]相一致。
2.3 不同遮阴处理对牡丹叶片光合速率日变化的影响
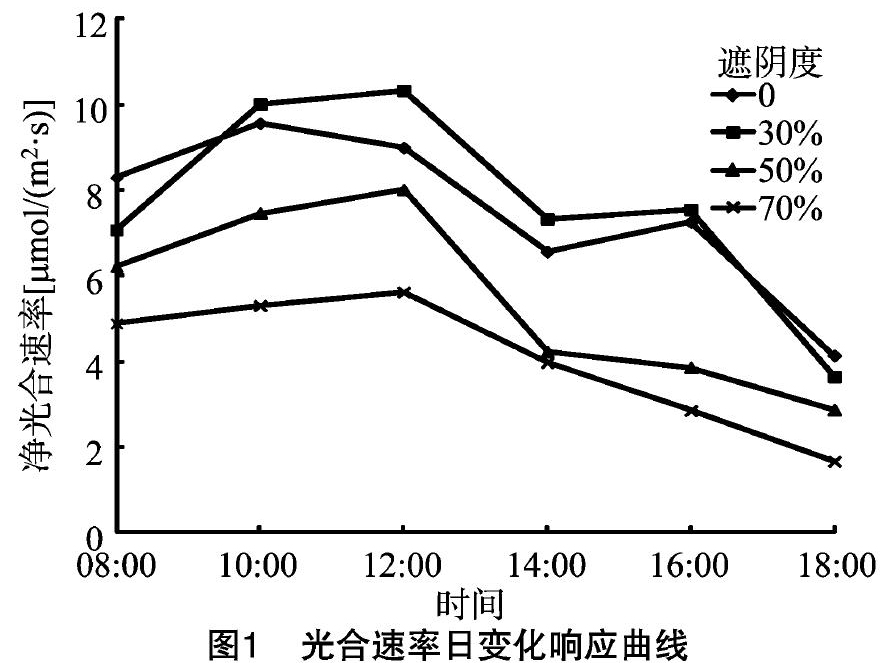
净光合速率是植物进行光合作用时积累干物质的重要基础和保障,在不同的季节和外部环境影响下会表现出很大的差异[18]。净光合速率曲线有3种表现类型,即“单峰”“双峰”和“三峰”。从图1可以看出,不同遮阴处理对牡丹净光合速率的影响差异较大,CK和30%遮阴度处理下净光合速率曲线为“双峰”型,出现了“光合午休”现象,而50%和70%遮阴度处理下,净光合速率曲线均为“单峰”型,无“光合午休”现象出现。早上日出后,随着光照辐射的增强,Pn值逐渐升高,未遮阴处理(CK)在10:00达到峰值,而不同遮阴处理下的峰值均出现在11:00至 12:00,与CK相比都有延迟。不同遮阴处理下,30%遮阴度处理的净光合速率峰值最高,达到 10.332 7 μmol/(m2·s),而70%遮阴度处理下的峰值最小,为5.618 8 μmol/(m2·s)。
从表3可以看出,随着遮阴度的增加,牡丹的净光合速率呈先升高后降低的趋势,而蒸腾速率、气孔导度和胞间CO2浓度均呈现出了递减趋势,可见遮阴处理对牡丹的光合各参数影响显著。在光照度降低的情况下,遮阴度为70%时牡丹叶片胞间CO2浓度和气孔导度急剧下降,与其他遮阴处理差异显著。胞间CO2浓度是影响植物光合作用的重要因素,而气孔又是直接影响水汽和CO2交换的重要通道,气孔过度关闭会对植物的蒸腾作用及土壤水分利用造成一定影响。
2.4 不同遮阴处理对牡丹光合响应的影响
采用非直角双曲线模型进行Pn-PAR曲线拟合,各处理拟合曲线的决定系数均大于0.99(表4)。在Pn-PAR响应曲线中(图2),在光合有效辐射<200 μmol/(m2·s)时,遮阴度为70%处理的叶片净光合速率明显高于其他遮阴处理。当光合有效辐射≥200 μmol/(m2·s)时,30%遮阴度处理的净光合速率明显高于其他遮阴处理。且净光合速率从高到低依次排列为T1>T2>CK>T3。
2.5 不同遮阴处理下光响应不同模型拟合结果对比
从表4可以看出,不同模型的曲线拟合程度存在较大差异,4种曲线拟合中直角双曲线的拟合度最低,而其他3种曲线的决定系数均大于0.98,拟合程度均优于直角双曲线模型。表4中数据显示,在CK、T1、T2、T3处理下,非直角双曲线模型拟合曲线的斜率分别为0.053 2、0.058 6、0.061 2、0.06 3,均小于相同遮阴度处理下其他模型拟合曲线的斜率,因此非直角双曲线模型拟合程度为最优。不同遮阴度处理下,指数方程模型拟合得到的暗呼吸速率在CK、T1、T2、T3处理下分别为1.079 5、1.073 6、1.069 2、1.066 4,均高于相同遮阴度处理下的其他模型,因此指数方程模型对暗呼吸速率(Rd)拟合程度为最优;同理,不同遮阴度处理下,指数方程模型拟合得到的光补偿点(Ic)也在相同遮阴度处理下最大,因此和其他模型相比,指数方程模型对光补偿点拟合程度为最优。
2.5.1 不同遮阴处理对光补偿点和光饱和点的影响 表4显示,随着遮阴程度的增加,牡丹叶片的光补偿点(Ic)和光饱和点(Isat)均有明显下降。不同植物的光补偿点差异较大,光补偿点越低,在弱光环境下进行光合作用的能力越强。在不同遮阴度环境下,叶片的光补偿点明显下降,说明其在弱光条件下对光能的利用能力较强。在重度遮阴环境下牡丹的净光合速率锐减,再加之胞间CO2浓度明显偏低(表3),说明气孔开放程度限制了CO2的输送[21]。
2.5.2 不同遮阴处理对表观量子效率的影响 光照度是植物进行光合作用的重要基础因素。表观量子效率能够有效反映出植物在生长过程中在不同环境下对光能的综合利用效率[22]。不同光照辐射梯度下的光合曲线斜率就体现了表观量子效率。曲线斜率越大,植物利用弱光的能力就越强;反之就越弱。表4显示,在非直角双曲线拟合下,70%遮阴度处理的斜率(α)为0.063 4,在不同遮阴度处理下为最高,说明其在弱光条件下对光能的利用能力较强[23]。
3 讨论与结论
光照是影响植物进行光合作用的重要外部环境因素,不同的光照度对植物光合作用和生长的影响是一个复杂漫长的过程[24]。光照条件对以观赏为主的花卉植物花期和开花数量、品质产生重要影响,对植物叶片的光合色素含量、光合响应和光合速率也会产生重要的影响。
随着遮阴度的增加,牡丹的物候期和花期有了明显延迟。70%遮阴度处理比无遮阴处理的盛花期延迟了7 d,在30%遮阴度处理下牡丹的开花数量和质量均达到了最佳。随着遮阴度的增加,叶绿素a、叶绿素b、总叶绿素含量均呈现增长趋势,叶绿素a/b的则呈减少趋势。
CK和30%遮阴度处理下牡丹净光合速率曲线为“双峰”型,出现了“光合午休”現象,而50%和70%遮阴处理下,均为“单峰”型。牡丹在未遮阴处理下10:00 Pn达到峰值,而不同遮阴处理下的峰值出现时间向后延迟。随着遮阴度的增加,牡丹的净光合速率呈先升高后降低的趋势,而蒸腾速率、气孔导度和胞间CO2浓度呈现出了递减趋势,可见遮阴处理对牡丹的光合参数影响显著。
在Pn-PAR响应曲线中,当光合有效辐射≥200 μmol/(m2·s)时,30%遮阴度处理的净光合速率明显高于其他遮阴处理。且净光合速率从高到低依次排列为CK>T1>T2>T3。在非直角双曲线拟合下,70%遮阴度处理下的表观量子效率最高,说明其在弱光条件下对光能的利用能力较强。植物的光补偿点和光饱和点被认为是评价植物耐阴性能的重要指标。光补偿点低且光饱和点高的植物适应周围环境能力强[25]。随着遮阴程度的增加,牡丹叶片的光补偿点(Ic)和光饱和点(Isat)均有明显下降。
参考文献:
[1]洪德元,潘开玉. 芍药属牡丹组的分类历史和分类处理[J]. 植物分类学报,1999,37(4):351.
[2]张凯月,贺春玲,侯小改,等. 油用牡丹经济价值和景观生态价值研究现状[J]. 中国农学通报,2019,35(10):66-71.
[3]尹宪志. 临夏气象[M]. 北京:气象出版社,2011.
[4]张新时,杨志杰,韩志刚. 几各光合作用响应典型模型的比较研究[J]. 武汉植物学研究,2009,27(2):197-203.
[5]刘 波,张庆费,夏 檑. 11种城市木本植物叶片生理物质对遮阴的响应[J]. 浙江林业科技,2007,27(6):15-18.
[6]苏文华,张光飞,李秀华,等. 光强和光质对灯盏花生长与总黄酮量影响的研究[J]. 中草药,2006,37(8):1244-1247.
[7]焦雨歆,赵 琦,王雪英,等. 环境因子对植物叶绿体结构的影响[J]. 生物技术通报,2008(2):5-10.
[8]Yamasaki S,Fujii N,Takahashi H. Photoperiodic regulation of CS-ACS2,CS-ACS4 and CS-ERS gene expression contributes to the femaleness of cucumber flowers through diurnal ethylene production under short-day conditions[J]. Plant,Cell & Environment,2003,26(4):537-546.
[9]李 程,裴忠孝,甘林叶,等. 光周期对春石斛开花及多胺含量的影响[J]. 植物生理学报,2014,50(8):1167-1170.
[10]王 静,罗 涛,梁 琴. 遮阴处理龙景春和太平红牡丹的光合生理和形态响应[J]. 贵州农业科学,2014,42(2):46-51.
[11]赵世杰. 植物生理学实验指导[M]. 北京:中国农业科学技术出版社,2002.
[12]Lichtenthaler H K,Wellburn A R.Determination of total carotenoids and chlorophylls a and b of leaf in different solvents[J]. Biochemical Society Transactions,1983,11:591-592.
[13]Miko U K,Graham D F. Investigation of the CO2 dependence of quantum yield and respiration and eucalyptus pauciflora[J]. Plant Physiology,1987,83(4):1032-1036.
[14]Marshall B,Biscoe P V.A model for C3 leaves describing the dependence of net photosynthesis on irradiance[J]. Journal of Experiment Botany,1980,120:29-39.
[15]Thornley J H M.Dynamic model of leaf photosynthesis with acclimation to light and nitrogen[J]. Annals of Botany,1998,81:431-436.
[16]叶子飘,于 强. 光合作用光响应模型的比较[J]. 植物生态学报,2008,32(6):1356-1361.
[17]Kupp M,Schulze E D. An empirical model of net photosynthesis and leaf conductance for the diurnal courses of CO2 and H2O exchange[J]. Journal of Plant Physiology,1985(12):513-526.
[18]周曙光,孔祥生,张妙霞,等. 遮光对牡丹光合及其他生理生化特性的影响[J]. 林业科学,2010,46(2):56-60.
[19]宋 洋,廖 亮,刘 涛,等. 不同遮阴水平下香榧苗期光合作用及氮分配的响应机制[J]. 林业科学,2016,52(5):55-63.
[20]安佰义,范爱淇,吴 双,等. 遮阴对玉带草光合特性的影响[J]. 江苏农业科学,2019,47(12):157-160.
[21]许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯,1997,33(4):241-244.
[22]杨志民,陈 煜,韩烈保,等. 不同光照强度对高羊茅形态和生理指标的影响[J]. 草业学报,2007,16(6):23-29.
[23]Lee D W,Done R A,Tarsis S L,et al. Correlates of leaf opticalproperties in tropical forest sun and extreme-shaded plants[J]. America Journal of Botany,1990,77(3):370-380.
[24]Powles S B. Phtoinhibition of photosynthesis induced by visible light[J]. Annual Review Plant Physiology,1984,35(35):15-50.
[25]余叔文. 植物生理與分子生物学[M]. 北京:科学出版社,1992.
- 工程监理工作中存在的弊端及优化策略分析
- 浅谈建筑工程造价的动态管理分析
- 浅谈园林施工管理策略及成本控制方法
- 浅析火力发电设备的营销策略
- 工程建设前期的造价控制
- 浅谈建筑工程招投标风险管理
- 浅析水利枢纽工程调度管理工作问题及对策
- 试析小型农村水利改革思路
- 智能电表用电监察工作开展思路
- 结合某项目做好低成本景观设计要点分析
- 基坑开挖对临近或下卧既有隧道的影响及研究进展
- GPS测量技术在工程测绘中的应用
- 邯郸市主城区道路交通拥堵分析与对策研究
- 水泥稳定碎石道路基层施工质量控制要点
- 水泥稳定碎石基层质量问题的处理及预防
- 建筑工程成本管理中BIM技术的运用
- 浅谈地铁地下线铺轨施工
- 红豆山隧道专项施工方案
- 浅析输变电线路施工的技术和措施
- 地铁给排水管材的选取与运用
- 浅谈加强水利工程施工管理的措施
- 铝模工程二次结构一次成型施工探索
- 浅析江肇高速公路塑料排水板软基处理方案
- 北方严寒地区城市轨道交通车型选择相关问题研究
- 浅谈电气工程安装施工中的要点
- retranslation
- retransmissions
- retransmission's
- retransmits
- retransmitted
- retransmitting
- retransmute
- retransplant
- retransplantation
- retransplantations
- retransplanted
- retransplanting
- retransplants
- retransport
- retransportation
- retraverse
- retraversed
- retraverses
- retraversing
- retreat
- retreatal
- retreated
- retreater
- retreaters
- retreating
- 沸腾的样子
- 沸腾纷乱的样子
- 沸腾翻滚
- 沸腾 融化
- 沸腾起来了
- 沸荡
- 沸跃
- 沸闹
- 沸鼎
- 沸鼎鱼游
- 油
- 油䯼髻上封官,粉鼻凹里受宣——裙带头衣食
- 油一路,水一路
- 油乎乎
- 油也得耗尽,明也看不见
- 油二代
- 油云
- 油井
- 油亮
- 油亮亮
- 油伙
- 油伞
- 油你涨
- 油儿
- 油儿拐