海力 肖瑶 庞文颖


摘要 为了研究Num1和Pil1之间是否存在互作,构建了Pil1超表达菌株,并通过酵母四分体解离技术成功构建得到用于免疫共沉淀试验的超表达Pil1和Num1的菌株。通过免疫共沉淀试验对Num1与Pil1的互作关系进行初步研究,并对试验中遇到问题的可能原因进行了分析。这些结果为研究Pil1是否参与细胞膜上Num1簇的形成奠定了基础。
关键词 酵母;Num1;Pil1;免疫共沉淀
中图分类号 Q78 ?文献标识码 A
文章编号 0517-6611(2020)18-0107-05
Abstract To study whether there is an interaction between Num1 and Pil1,we constructed a Pil1 overexpression strain,and then successfully constructed the strain overexpressing both Pil1 and Num1,which can be used for coimmunoprecipitation experiments,via yeast tetrad dissection technique.We conducted preliminary study on the interaction between Num1 and Pil1 by coimmunoprecipitation,and analyzed possible reasons for problems encountered in the experiment.These results laid the foundation for studying whether Pil1 participates in the formation of Num1 clusters on the cell membrane.
Key words Yeast;Num1;Pil1;Coimmunoprecipitation
細胞器(organelle)一般认为是散布在细胞质内具有一定形态和功能的微结构或微器官。细胞中的细胞器主要有线粒体、内质网、中心体、叶绿体、高尔基体、核糖体等。它们组成了细胞的基本结构,使细胞能正常工作、运转。曾经认为不同的细胞器在细胞中执行不同的功能,各细胞器独立运作,但现在发现,细胞器之间是相互联系的,并且这些联系对于细胞器的功能和整个细胞内环境的稳定至关重要[1-3]。
线粒体与质膜间的接触对于细胞正常生长也很重要,这些接触会对线粒体形态、分裂、融合、定位和遗传进行调控[4],线粒体与质膜的连接使得线粒体附着在细胞皮层,使线粒体保持延伸状的形态,更重要的是这种连接会确保在有丝分裂过程中线粒体被准确地分配至子细胞中[5],同时,这种线粒体-质膜连接也参与到线粒体分裂[6-7]。
细胞器间的接触是通过细胞器间的蛋白互作实现。并且,在不同的生理环境下定位于接触位点的蛋白质会建立、维持或者改变接触,从而影响多种细胞的基本功能[8]。在酿酒酵母中,线粒体-内质网(ER)-质膜(PM)三者间形成的锚结构(MECA)将线粒体与PM和ER相连,使得3个细胞器膜彼此靠近[9], MECA的核心蛋白组分是Num1蛋白,Num1是一个313 kD蛋白,由一个N末端卷曲螺旋(CC)结构域(95~303 aa),一个推测的钙结合区EF-hand(303~316 aa),一个FFAT基序(306~330 aa),一个包含13个由64个残基串联重复序列构成的(592~1 776 aa)的中间TR结构域,以及一个C末端的PH(pleckstrin homology)结构域组成[10-11]。Num1通过C末端的PH结构域与质膜结合,通过N末端的CC结构域与线粒体结合,从而使线粒体和细胞质膜连接起来。有趣的是,线粒体与质膜之间的Num1蛋白簇也作为动力蛋白的皮质锚,在纺锤体的正确定位中起重要作用[12-14]。动力蛋白沿着星状微管运行,之后被锚定至细胞膜的Num1簇[15],以帮助有丝分裂过程中纺锤体的正确定位[12-13]。反过来,Num1簇的形成也受到线粒体的调控,当线粒体遗传受到抑制时, Num1簇的形成受到影响,进而导致动力蛋白介导的纺锤体定位产生错误,导致细胞核的不正常分离[16]。
质膜是否也在Num1簇的形成中起作用,为了研究这一问题,从实验室之前通过Num1的Pull-Down试验鉴定出的蛋白中,筛选出定位在细胞膜的蛋白Pil1进一步研究[17]。Pil1是酵母Eisosome的主要成分,Eisosome是一种大的蛋白复合体,定位于酵母质膜上[18]。Eisosome通过Pil1直接与细胞膜上的脂质结合并产生内陷[19]。为了验证Pil1和Num1之间的互作,构建了过表达 Pil1 和 Num1 的菌株,通过免疫共沉淀试验检测 Pil1 和 Num1 的相互作用,旨在为研究Eisosome是否在Num1簇的形成中起作用奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株。 酿酒酵母YWL37基因型:MATα ura3-52 lys2-801 leu2-Δ1 his3-Δ200 trp1-Δ63[17];YT52基因型:MATa TRP1-GAL1-Num1-YFP-HIS ura3-52 lys2-801 leu2-Δ1 his3-Δ200 trp1-Δ63;质粒pFA6a- TRP-GAL1-3HA;免疫共沉淀试验所需菌株收集量为YT47:135 mg;YT52:166 mg;YT84:149 mg。
1.1.2 培养基及试剂。YPD培养基和缺乏相应氨基酸的SD (synthetic define)培养基;SPM培养基(Yeast extract 1 g/L,CSM 0.2 g/L,Glucose 0.5 g/L,Potassium acetate 10 g/L);Lithium-Sorb溶液(10 mmol/L Trisbase,100 mmol/L Sorbitol,10 mmol/L LioAC,1 mmol/L EDTA,pH 8.0);PEG(40% PEG,10 mmol/L LioAC,10 mmol/L Trisbase,1 mmol/L EDTA,pH 8.0);转膜液(10%甲醇,0.05%SDS,25 mmol/L Trisbase,190 mmol/L Glycine);TBST溶液(50 mmol/L Trisbase,300 mmol/L NaCl,0.05% Tween 20,pH 8.0)。
1.2 方法
1.2.1 GAL1-3HA-PIL1菌株的构建。利用聚合酶链式反应(PCR) 对PIL1进行过表达启动子和蛋白标签的标记。
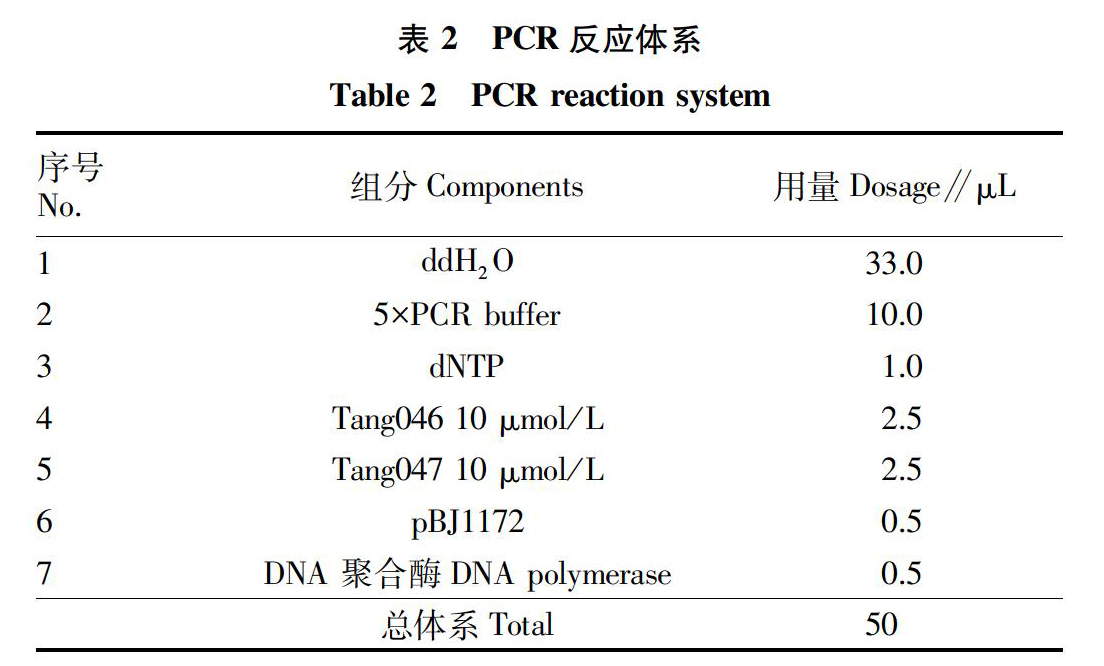
1.2.1.1 PCR扩增转化片段。为了后续更好地筛选到Pil1高表达量菌株,以pFA6a-TRP-GAL1-3HA质粒为模板扩增了标记PIL1的转化片段TRP-GAL1-3HA。引物序列和PCR反应条件见表1、2。
PCR扩增程序:第一步98 ℃ 预变性1 min;第二步98 ℃ 变性10 s,45 ℃ 退火20 s,72 ℃ 延伸1 min(根据PCR预计产物片段长度调整),2个循环;第三步98 ℃ 变性10 s,53 ℃ 退火20 s,72 ℃ 延伸1 min,33个循环;第四步72 ℃ 彻底延伸 8 min,最后 12 ℃ 保存。
PCR扩增完成之后每50 μL PCR反应产物加 5 μL 5×loading buffer混匀后进行琼脂糖凝胶电泳检测,电泳检测后用凝胶提取试剂盒回收,将回收产物放至4 ℃ 暂存。
1.2.1.2 酵母感受态细胞制备。从-80 ℃ 冰箱中取所需菌株在YPD固体培养基上划线,放置30 ℃ 恒温培养箱培养1~2 d,挑菌落到3 mL YPD液体培养基,30 ℃,220 r/min 摇床培养24 h左右;次日吸取6 μL菌液(视酵母菌长势而定)到新的30 mL YPD液体培养基,30 ℃、220 r/min 摇床培养至对数期 (OD600 约为0.5);3 000 r/min 离心3 min弃上清,收集30 mL 对数期生长的YWL37细胞,取10 mL 灭菌水重悬清洗细胞,之后3 000 r/min 离心1 min去上清,加3 mL Lithium-Sorb溶液吹打重悬细胞,3 000 r/min 离心2 min 去上清;用360 μL Lithium-Sorb 和40 μL 鲑鱼精子DNA (作为载体DNA,使用之前应先在沸水中煮10 min 后放冰上冷却) 重悬浮细胞,分装到新的1.5 mL离心管中,每管100 μL,于-80 ℃ 保存。
1.2.1.3 目的片段转化。取100 μL YWL37感受态细胞置于冰上;取10 μL PCR回收产物TRP-GAL1-3HA至100 μL YWL37感受态细胞中,加入6倍体积PEG混匀,室温静置30 min;然后加入38 μL 二甲基亚砜(DMSO),42 ℃ 水浴热激5 min,之后3 000 r/min 离心2 min,弃上清;加50 μL 灭菌水重悬细胞,之后将细胞涂布缺乏色氨酸的固体培养基SD-TRP。
1.2.1.4 阳性克隆的筛选。在对应的SD-TRP培养基上挑选单克隆,在新的SD-TRP培养基上划线筛选单克隆,重复1次;将从第二次划线的培养基上挑选的单克隆进行扩大培养,待提酵母总蛋白进行鉴定。
1.2.2 重组转化子的过表达鉴定。
1.2.2.1 GAL1启动子超表达菌株细胞培养。将转化筛选得到的克隆在YPD固体培养基划线,30 ℃恒温24 h,之后接种细胞至3 mL YPD液体培养基,30 ℃、220 r/min 摇床中培养至饱和(约24 h)。取6 μL生长饱和的细胞液,转移至3 mL含有2%棉子糖(Raffinose)的YPA液体培养基中,30 ℃、220 r/min 摇床中培养16 h。将培养16 h的细胞液,3 000 r/min 离心2 min,弃上清,向沉淀中加入含2%半乳糖(Galactose)的YPA液体培养基,30 ℃、220 r/min摇床中继续培养4 h,之后收集细胞。
1.2.2.2 TCA法制备酵母总蛋白。3 000 r/min离心5 min收集上述培養的细胞至收集管,向沉淀中加入100 μL 20%三氯乙酸(TCA) ,之后加直径1 mmol/L玻璃珠至液面以下,室温使用珠式细胞破碎仪(Minibeeadbeater16,Biosepc)破碎细胞1 min,冰上冷却2 min,重复3次。用针头在收集管底部扎一小孔,将收集管放入1.5 mL离心管中,4 ℃ 1 000 r/min离心3 min,弃收集管,3 000 r/min继续离心5 min,弃上清。向沉淀中加40 μL 2×蛋白上样缓冲液(protein loading buffer) 和40 μL Tris-HCl(pH 8.0)混匀,之后沸水中煮5 min,12 000 r/min 离心1 min,-20 ℃保存,上清用于蛋白质免疫印迹。
1.2.2.3 蛋白免疫印迹。该试验共检测2种蛋白:Pil1和Num1。分别采用10%变性聚丙烯酰胺凝胶及6.25%变性聚丙烯酰胺凝胶电泳,SDS-PAGE胶配方见表3、4。电泳结束后,针对Pil1使用半干转仪在含10%甲醇的转膜液中将蛋白转移至PVDF膜,针对Num1则使用湿转的办法在相同的转膜液中将蛋白转至PVDF膜。将PVDF膜浸泡至含5%脱脂奶粉的TBST溶液轻摇1 h。封闭之后,使用HA抗体(THETMc-HA Tag Antibody,mAb,Mouse,GenScript)或GFP抗体(Mouse Anti GFP Monoclonal-Antibody,兴华基因)室温轻摇孵育2 h,TBST溶液洗3次,每次10 min,之后使用二抗室温轻摇孵育1 h,TBST溶液洗3次,每次10 min,然后使用含3 μL H2O2 的10 mL显色液对PVDF膜上的蛋白进行显色,凝胶成像仪曝光检测。
Western Blot检测显示Pil1成功超表达,命名为YT47 (GAL1-TRP-3HA-PIL1)。
1.2.3 酵母四分体解离法构建免疫共沉淀(CoIP)菌株及表达鉴定。
1.2.3.1 亲代酵母菌株杂交。取YT47、YT52在YPD固体培养基上划线,30 ℃恒温培养1~2 d,长出细胞后,挑取适量的亲代菌株在新的YPD固体培养基上混合,30 ℃恒温培养24 h,用牙签挑混合菌落顶部少量菌在相应双缺培养基SD-TRP-HIS上划线培养1~2 d,之后挑取菌落转移至3 mL SPM液体培养基中,30 ℃、220 r/min培养3~4 d(不能超过4 d),普通光学显微镜下观察是否有四分体存在。
1.2.3.2 酵母细胞壁去除。取70 μL SPM中细胞液至1.5 mL离心管,3 000 r/min离心1 min,弃上清。加100 μL Lithium-Sorb溶液和8 μL Zymolase混匀,37 ℃静置8 min,之后进行四分体解离或存放于4 ℃直至使用。
1.2.3.3 酵母四分体解离及菌株筛选。取细胞沉淀20 μL滴至YPD固体培养基,竖立培养皿,使沉淀在培养基表面成一条直线,利用酵母四分体解离操作仪(MSM400,Singer)按照使用说明书,进行四分体分离,解离完成后将培养基放置30 ℃恒温培养箱培养至细胞长出,之后根据需要得到相应的菌株,将培养皿复制至SD-HIS,SD-TRP培养基和交配型选择培养基培养,筛选结果如表5。
1.2.3.4 Western Blot鉴定。将筛选得到的菌落转移至 3 mL YPD 液体培养基中培养,收集对数期细胞提取总蛋白并进行 Western Blot 鉴定。
1.2.4 CoIP检测Pil1与Num1的互作。
收集细胞:每个CoIP需要用不少于90 mg的细胞。
细胞培养:第1天,从-80 ℃冰箱中取出CoIP所需菌株,用YPD培养基活化;第3天早上从平板上取细胞接种到3 mL YPD液体培养基中;第4天,等细胞长至饱和,下午吸取108 μL 菌液至54 mL 液体培养基中(48.6 mL YPA-D+5.4 mL 20% Raffinose)过夜培养16 h;第5天,3 000 r/min离心5 min,弃上清,加入54 mL YPA+半乳糖培养4 h,收集细胞。
免疫共沉淀试验方法:
(1)细胞长好以后,3 000 r/min离心2 min弃上清,再加入1 mL灭菌水重悬,3 000 r/min离心1 min弃上清,加入1 mL 1× 细胞裂解液(lysis buffer)[配方见表6、7,不含蛋白酶抑制剂(protease inhibitor cocktail),购自Roche]重悬后转入1.5 mL收集管中,3 000 r/min离心1 min弃上清后可在-80 ℃保存。
(2) 吸取30 μL Protein G磁珠(GE Healthcare)到1.5 mL 离心管(EP管)中(剪枪头),加入1 mL 1×lysis buffer(不含PI)清洗,之后将EP管放入磁力架MagRack 6,收集磁珠到离心管一侧,吸弃上清,再加入1 mL 1× lysis buffer(不含PI)重复洗一次弃上清,加入200 μL 1×lysis buffer(含PI)洗一次弃上清后,向EP管中加入200 μL 1×lysis buffer和3 μL GFP抗体,4 ℃孵育。
(3)细胞破碎:向已收集好的细胞中(保存在-80 ℃的细胞取出后需置于冰上融化)先加入300 μL 1×lysis buffer(含PI),重悬后加入1×lysis buffer(含PI)至600 μL,然后向收集管中加入珠子至液面以下。细胞破碎45 s,拿出放置在冰上静置2 min,重复3个循环。
(4)在收集管底部戳一个洞,插到1.5 mL EP管中,4 ℃、3 000 r/min离心2 min后,向收集管中加入200 μL 1×lysis buffer(含PI),继续3 000 r/min离心2 min,弃收集管,再12 000 r/min离心15 min,取50 μL上清作为Input。
(5)吸取剩余上清(大约有600 μL)到孵育抗体的1.5 mL EP管中,4 ℃继续孵育1.5 h。
(6)孵育完后,用磁力架收集磁珠,取50 μL上清作为未结合蛋白(unbound)后棄上清。
(7)用1×洗脱液(wash buffer)(不含PI) 洗磁珠3次,每次加1 mL 1× wash buffer。
(8)用磁力架收集磁珠到离心管一侧,弃上清后加入40 μL ddH2O和40 μL 2× protein loading buffer, 沸水煮5 min,-20 ℃保存。 向Input和unbound中加入50 μL 2× protein loading buffer沸水煮5 min,-20 ℃保存。
(9)点样前拿出样品沸水煮5 min,短暂离心,取上清点样,进行Western Blot检测。
2 结果与分析
2.1 GAL-3HA-Pil1重组菌株的构建和表达鉴定 为了检测Pil1与Num1在细胞内是否发生互作,以质粒pFA6a-TRP-GAL1-3HA为模板扩增TRP1-GAL1-3HA片段(1 798 bp)(图1),并将此片段转化至YWL37野生型菌株,通过Western?Blot检测到Pil1的表达(图2),构建得到了TRP1-GAL1-3HA-PIL1超表达菌株,命名为YT47。
2.2 酵母四分体解离法构建重组菌株 由于用于酵母杂交的亲代酵母中均带有TPR标记,所以根据酵母四分体筛选分离后再用Western Blot进一步鉴定(图3),筛选得到A2(MATa TRP-GAL-3HA-PIL1 TRP-GAL1-NUM1-YFP-HIS 命名为YT84),B1(MATa TRP-GAL-3HA-PIL1 TRP-GAL1-NUM1-YFP-HIS 命名为YT85)。
HA抗体检测A2、B1菌株中Pil1是否超表达。结果显示A2、B1菌株构建成功,将用于免疫共沉淀(CoIP)试验。
2.3 CoIP检测Num1与Pil1的互作 CoIP试验使用GFP抗体(兔抗)(Genescript)孵育Protein G,经GFP抗体沉淀下来的蛋白分别用GFP抗体和HA抗体检测,试验结果如图4所示。Num1-GFP蛋白被成功地沉淀下来(图4上右,第2道和第3道)。而且,3HA-Pil1蛋白也随着Num1-GFP蛋白共同沉淀下来(图4下右,第3道)。但是,在不表达Num1-GFP而只表达3HA-Pil1 的CoIP样品中也检测到3HA-Pil1蛋白(图4下右,第1道),说明3HA-Pil1蛋白与免疫沉淀用的GFP抗体或Protein磁珠发生了非特异性结合。
3 結论与讨论
该研究通过基因同源重组及酵母四分体解离等方法成功构建了TRP-GAL-3HA-PIL1 TRP-GAL1-NUM1-YFP-HIS菌株,旨在通过免疫共沉淀试验探究Num1与Pil1的互作关系,以期为后续研究Pil1所在的Eisosome膜结构是否影响细胞膜上Num1簇(Num1多聚体)的形成奠定基础。在用Protein G磁珠和GFP抗体进行的CoIP试验中,虽然3HA-Pil1能和Num1-GFP共同沉淀下来,但是在只表达3HA-Pil1而不表达Num1-GFP的细胞沉淀下来的样品中也检测到了3HA-Pil1。分析这种非特异性条带的出现可能与以下几方面有关:①Protein G磁珠能非特异性结合3HA-Pil1蛋白;②用于免疫沉淀试验的GFP抗体能非特异性结合HA标记的Pil1蛋白,使其沉淀下来;③免疫共沉淀试验中的清洗步骤不彻底,使得Pil1与Protein G间产生非特异性粘黏。
为得到无干扰的试验结果,为后期试验奠定基础,可以从以下3个方面进行改善:①更换不同品牌的Protein G;②检测GFP 抗体的特异性;③调整免疫共沉淀试验中lysis buffer的盐离子浓度,提高清洗的严谨性。
参考文献
[1] BRAUN R J,WESTERMANN B.With the help of MOM: Mitochondrial contributions to cellular quality control[J].Trends in cell biology,2017,27 (6):441-452.
[2] DENIC V,DTSCH V,SINNING I.Endoplasmic reticulum targeting and insertion of tailanchored membrane proteins by the GET pathway[J].Cold spring harbor perspectives in biology,2013,5(8):1-11.
[3] DYALL S D,BROWN M T,JOHNSON P J.Ancient invasions:From endosymbionts to organelles[J].Science,2004,304(5668):253-257.
[4] KLECKER T,WEMMER M,HAAG M,et al.Interaction of MDM33 with mitochondrial inner membrane homeostasis pathways in yeast[J].Scientific reports,2016,5(1):1-14.
[5] YU J W,MENDROLA J M,AUDHYA A,et al.Genomewide analysis of membrane targeting by S.cerevisiae pleckstrin homology domains[J].Molecular cell,2004,13(5):677-688.
[6] HEILCHAPDELAINE R A,OBERLE J R,COOPER J A.The cortical protein Num1p is essential for dyneindependent interactions of microtubules with the cortex[J].The journal of cell biology,2000,151(6):1337-1343.
[7] FARKASOVSKY M,KNTZEL H.Cortical Num1p interacts with the dynein intermediate chain Pac11p and cytoplasmic microtubules in budding yeast[J].The journal of cell biology,2001,152(2):251-262.
[8] LABROUSSE A M,ZAPPATERRA M D,RUBE D A,et al.C.elegans dynaminrelated protein DRP1 controls severing of the mitochondrial outer membrane[J].Molecular cell,1999,4(5):815-826.
[9] LACKNER L L,PING H,GRAEF M,et al.Endoplasmic reticulumassociated mitochondriacortex tether functions in the distribution and inheritance of mitochondria[J].Proceedings of the national academy of sciences,2013,110(6):E458-E467.
[10] INGERMAN E,PERKINS E M,MARINO M,et al.Dnm1 forms spirals that are structurally tailored to fit mitochondria[J].The journal of cell biology,2005,170(7):1021-1027.
[11] GREENBERG S R,TAN W M,LEE W L.Num1 versus NuMA:Insights from two functionally homologous proteins[J].Biophysical reviews,2018,10(6):1631-1636.
[12] KNOBLACH B,RACHUBINSKI R A.Sharing the cells bountyorganelle inheritance in yeast[J].Journal of cell science,2015,128(4):621-630.
[13] TANG X,GERMAIN B S,LEE W L.A novel patch assembly domain in Num1 mediates dynein anchoring at the cortex during spindle positioning[J].The journal of cell biology,2012,196(6):743-756.
[14] LAWRENCE E J,MANDATO C A.Mitochondria localize to the cleavage furrow in mammalian cytokinesis[J].PLoS One,2013,8(8):1-12.
[15] FRIEDEN M,ARNAUDEAU S,CASTELBOU C,et al.Subplasmalemmal mitochondria modulate the activity of plasma membrane Ca2+ATPases[J].Journal of biological chemistry,2005,280(52):43198-43208.
[16] LACKNER L L.The expanding and unexpected functions of mitochondria contact sites[J].Trends in cell biology,2019,29(7):580-590.
[17] 唐仙英,肖瑤,海力.蛋白体外结合实验联合质谱分析鉴定Num1的互作蛋白[J].中南民族大学学报(自然科学版),2018,37(4):27-30.
[18] MOREIRA K E,WALTHER T C,AGUILAR P S,et al.Pil1 controls eisosome biogenesis[J].Molecular biology of the cell,2008,20(3):809-818.
[19] KABECHE R,BALDISSARD S,HAMMOND J,et al.The filamentforming protein Pil1 assembles linear eisosomes in fission yeast[J].Molecular biology of the cell,2011,22(21):4059-4067.
- 医务社工介入贫困患儿救助的资源分析
- 从比较法看童工保护
- 热点事件报道中的城市形象建构及传播策略
- 捕捞渔民对海洋环境变迁的主观感受
- 浅析我国建国后婚姻自由的现状与问题
- 中西同性恋法律地位变化解析
- 基于劳动法视域中劳动关系认定问题的思考
- 探讨社会保障法与劳动法的界定
- 浅析受害人近亲属缺位的侵权死亡赔偿
- 网络侵权行为下对公民安宁权的诉求
- 商标许可增值利益的归属与分配
- 论链接服务侵犯著作权中的服务器原则适用
- 手机游戏的著作权保护路径探究
- 论网络著作权的侵权与保护
- 河南省科技型中小企业知识产权运用能力提升研究
- 物权法下宅基地使用权的转让
- 法律视角下的女性犯罪原因及预防对策
- 论我国共犯性质的现状与出路
- 武警部队遂行救援任务法律保障初探
- 办理盗窃犯罪适用新司法解释过程中的难点及探讨
- 入户抢劫的理解与适用难度探析
- 工程建设领域职务犯罪的易发环节和预防对策研究
- 自洗钱行为在我国不独立定罪的缘由分析
- 浅析销售假冒注册商标的商品罪的特点、侦办难点及打防对策
- “口袋罪”
- dislikes
- dislike²
- dislike¹
- disliking
- dislimb
- dislink
- disload
- dislocate
- dislocated
- dislocatedly
- dislocates
- dislocating
- dislocation
- dislocation's
- dislocative
- dislocatory
- dislodge
- dislodgeable
- dislodged
- dislodg(e)ment
- dislodgements
- dislodges
- dislodging
- dislodgments
- disloyal
- 强风造成的灾害
- 强食
- 强食弱肉
- 强饭
- 强饭廉颇
- 强饭自爱
- 强饮强食
- 强鲠峭立
- 强黠
- 强龙不压地头蛇
- 强龙不压地头蛇。
- 强龙压不住地头蛇
- 强龙斗不过地头蛇
- 强龙难敌地头蛇
- 强:
- 弼
- 弼亮
- 弼佐
- 弼匡
- 弼导
- 弼直
- 弼谐
- 弼辅
- 弼违
- 弼非