王方雨 张运尚 胡曼 樊剑鸣

摘要 为了制备高灵敏度、高特异性的抗新霉素单链抗体,试验采用从杂交瘤细胞中提取单链抗体基因,转入大肠杆菌表达系统进行表达。然后采用分子对接的方法研究抗新霉素单链抗体和新霉素分子的识别机制并进行虚拟突变,将关键结合位点的氨基酸残基进行突变,在对突变体进行表达和纯化后建立间接竞争酶联免疫吸附分析方法。结果表明:氨基酸残基Leu227为单链抗体与新霉素结合的关键氨基酸残基,将其突变为Asp,获得突变体。IC 50为1.58 ng/mL,相比于亲本单链抗体的26.23 ng/mL,突变体的敏感性提高了16.6倍。试验成功制备了高灵敏度、高特异性的抗新霉素单链抗体,为动物源性食品中的抗生素残留检测提供了研究基础。
关键词 单链抗体;氨基糖苷类抗生素;药物残留检测;分子识别机制;间接竞争酶联免疫吸附法
中图分类号 Q789? 文献标识码 A
文章编号 0517-6611(2020)20-0103-05
doi:10.3969/j.issn.0517-6611.2020.20.028
Preparation and Evolution of Anti-Neomycin Single-chain Antibody Based on Molecular Docking Technology
WANG Fang-yu1,ZHANG Yun-shang 1,2,HU Man1 et al (1.Key Laboratory of Animal Immunology,Henan Academy of Agriculture Sciences,Zhengzhou,Henan? 450002;2.College of Public Health, Zhengzhou University,Zhengzhou,Henan 450001)
Abstract In order to prepare high-sensitivity and high-specificity anti-neomycin single-chain antibodies, the experiment used genes extracted from hybridoma cells and directly transferred to E.coli expression system for expression.Then, the molecular docking method was used to study the recognition mechanism of anti-neomycin single-chain antibody and neomycin molecule and virtual mutation was performed.The amino acid residues of the key binding site were mutated, and the indirect competitive enzyme-linked immunosorbent assay method was established after expression and purification of the mutant.The results showed that the amino acid residue Leu227 was the key amino acid residue for the binding of single-chain antibody to neomycin, and it was mutated to Asp to obtain a mutant.The IC 50 was 1.58 ng/mL, which was 16.6 times more sensitive than the parental single-chain antibody (26.23 ng/mL).It showed that the test successfully prepared high-sensitivity and high-specificity anti-neomycin single-chain antibody,which provided a research basis for the detection of antibiotic residues in animal-derived foods.
Key words Single-chain antibody;Aminoglycosides;Drug residue detection;Molecular recognition mechanism;Indirect competitive enzyme-linked immunosorbent assay
新霉素(neomycin, NEO)是一种氨基糖苷类抗生素(aminoglycosides, AGs),该类药物因含有多个氨基和羟基基团,所以对革兰氏阳性菌和阴性菌都有抗菌作用,同时还具有极性强、水溶性好,在碱性环境抗菌性强等诸多优点 [1]。作为广谱抗菌剂和促进动物生长发育的饲料添加剂 [2],新霉素被广泛应用在畜牧业中。养殖人員不合理或不规范的使用可能会导致养殖动物体内中残留抗生素,并产生细菌耐药性 [3],当体内有药物残留的动物被加工为肉类上市时,人们食用后会对人体造成多种危害,主要包括肾毒性、耳毒性和神经肌肉阻滞等 [4]。为了防止动物性食品中氨基糖苷类药物残留对公众健康构成威胁,欧盟、粮农组织/世卫组织食品添加剂联合专家委员会和中国制定了可食用食品中NEO的最高残留限量 [5]。如在牛奶中新霉素的MRL为200 μg/kg,在鸡肉组织中为100 μg/kg。
鉴于抗生素在环境和食物链中的广泛传播,其对生态环境和人类健康的不利影响受到广泛关注。目前已有众多方法对食物样品进行检测 [6-8],如超高效液相色谱法、液相色 谱-质谱法和免疫法。其中基于抗体与抗原特异性结合原理的酶联免疫吸附试验(ELISA)被认为是检测抗体最快速、简便的方法 [9-11],适用于对样品的初步筛选。在以往的研究中,研究人员已经开发了基于单克隆抗体(MAb)的间接竞争酶联免疫吸附法(IC-ELISA) [12-13]。但传统抗体单抗或多抗的制备均较为复杂,不仅浪费时间和精力,还有不可进化的弊端 [3]。有必要开发一种简单、快速、高效的新型抗体。目前,单链抗体(ScFv)是最受关注的抗体之一。ScFv是通过将抗体的重链可变区(VH)和轻链可变区(VL)通过接头(Linker)连接而成的抗体。不仅可以通过分子对接技术改善其识别机制,还可以进行随机诱变和定点诱变进行改造和进化 [14]。在这项研究中,为了获得高灵敏度的抗NEO-ScFv,从能分泌抗NEO单克隆抗体的杂交瘤细胞中提取重链和轻链的基因,构建ScFv基因,然后转染到大肠杆菌BL21(DE3)中进行表达。通过分子对接方法研究其识别机制,定向诱变进化了ScFv。获得灵敏度更高的ScFv突变体,并以此建立IC-ELISA检测方法,以期为检测动物性食品中新霉素残留奠定了 基础。
1 材料与方法
1.1 材料 抗NEO单克隆抗体杂交瘤细胞由河南省农业科学院动物免疫学重点实验室提供。
1.2 药品
新霉素標准品、庆大霉素标准品、链霉素标准品和卡那霉素标准品均购于中国兽医药品监察所,药物均为分析纯或更高。
1.3 主要试剂
TRIzol总RNA提取试剂盒、PrimeScript RT-PCR试剂盒购自美国Invitrogen公司;羊抗鼠酶标二抗购自北京Solarbio公司;AEC酶底物试剂盒购自北京ZSGB-bio公司;PBST溶液由包含0.05%Tween-20的PBS配制;异丙基-β-D-硫代半乳糖苷(IPTG),限制性内切酶(Sfi Ⅰ和Not Ⅰ)和T4 DNA连接酶购自TaKaRa公司(中国,大连);EasyPure快速凝胶提取试剂盒,EasyPure质粒MiniPrep试剂盒,表达载体Pet-32a感受态细胞Rosetta-gami(DE3),快速多位点诱变系统和Luria-Bertani培养基(液体和固体)购自TransGen生物公司(中国,北京);DNA纯化试剂盒和SDS-PAGE凝胶制备试剂盒来自北京ComWin生物技术有限公司(中国,北京);蛋白胨、酵母提取物购自英国Oxoid公司;引物的合成和基因序列分析在Sangon生物公司(中国,上海)进行。
1.4 研究方法
1.4.1 重链、轻链基因的扩增与组装。
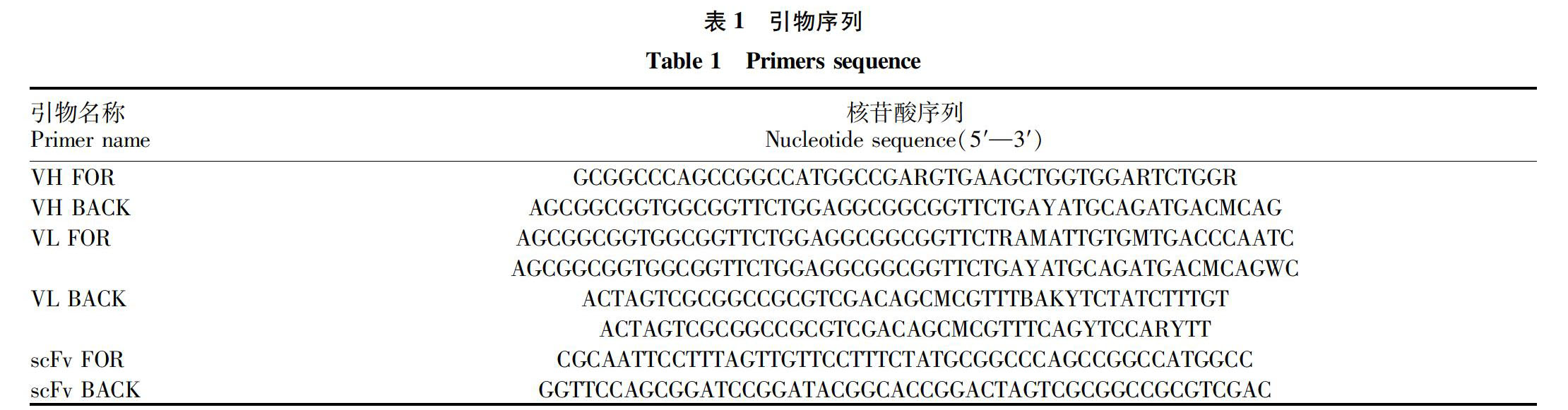
使用TRIzol总RNA提取试剂盒从抗NEO杂交瘤细胞中提取总RNA。然后,以总RNA为模板使用PrimeScript RT-PCR试剂盒进行反转录合成cDNA。使用VH和VL引物扩增的全套ScFv基因,引物序列见表1。反应条件如下:94 ℃ 预变性5 min;94 ℃持续45 s,58 ℃持续60 s,72 ℃持续45 s,共进行30个循环;最后在72 ℃下延伸10 min。反应产物经琼脂糖凝胶电泳鉴定,用EasyPure快速凝胶提取试剂盒回收目的片段。将获得的VH和VL片段分别与载体连接,进行基因序列分析。然后以编码肽(Gly4Ser)3为接头,将VH和VL基因通过重叠延伸PCR(SOE-PCR)拼接为完整的ScFv基因。反应条件为94 ℃ 5 min;94 ℃ 45 s,68 ℃ 60 s,72 ℃ 45 s,共进行30个循环;最终在72 ℃下延伸10 min。PCR产物通过琼脂糖凝胶电泳验证,并对相关片段进行测序。
1.4.2 抗NEO单链抗体的测序、表达和纯化。
利用Ncol Ⅰ和Not Ⅰ限制性酶消化靶基因和原核表达载体pET-32a,并通过T4 DNA连接酶进行连接。然后将重组质粒转化到大肠杆菌(DE3)中,并放置在37 ℃含有100 μg/mL卡那霉素的Luria-Bertani(LB)培养基中培养。细菌溶液的OD 600达0.6~0.8后,将1 mmol/L IPTG加入培养物中以诱导ScFv的表达,然后使培养物在37 ℃进一步生长16 h。收集上清液,并使用MWCO:8 000~14 000 Da透析袋在PEG/NaCl中将其浓缩至100倍。将收集的沉淀物分别用PBS重悬以获得周质裂解物,并通过超声处理裂解以获得全细胞裂解物。上清液、周质裂解物和全细胞裂解物用于通过SDS-PAGE分析蛋白质的溶解度。最后,使用Beaver Bead TM His-tag蛋白纯化磁珠纯化抗NEO ScFv蛋白。
1.4.3 同源性建模和分子对接。
首先根据测序结果,在NCBI数据库(https:∥www.ncbi.nlm.nih.gov/)中搜索了抗NEO单链抗体模型的可能模板序列,并在Blast进行了序列比较,选择模板与抗NEO单链抗体模型具有高度一致性。具有高得分和低e值的序列被用作抗NEO单链抗体模型构建的模板序列。然后,利用SWISS MODEL在线服务器建立抗NEO ScFv的模型。为了验证同源性建模结果的可靠性并确定最佳模型结构,使用Procheck、Verify3D和ERRAT程序评估了构建的抗NEO单链抗体同源性模型的一致性,并选择了最佳受体模型用于进一步的分子对接研究。使用MOE 2015.10进行分子对接,分析NEO与抗NEO-ScFv的结合模式并找到关键残基。在Dock模块中,通过诱导拟合和在Amber10:EHT力场下,将NEO停靠在抗NEO-ScFv的活性位点中。使用默认参数后具有30个对接构象的对接配体用于进一步分析。
1.4.4 ScFv抗体的定向诱变。
基于抗NEO-ScFv与NEO的结合模式研究分析,通过虚拟突变单链抗体与NEO结合关键残基,可以提高NEO与单链抗体的结合亲和力。利用MOE 2015.10软件进行单链抗体氨基酸残基的虚拟突变,关键结合氨基酸残基Leu227被Asp代替来产生抗NEO-ScFv的突变体。然后,优化了虚拟突变ScFv突变模型的结构,获得了稳定的ScFv突变模型。随后,通过诱导拟合的方法对具有ScFv突变的NEO进行对接研究,并使用默认参数获得30个构象。在试验过程中,将表达载体ScFv-pCANTAB5E中的ScFv基因进行突变,以利用快速多位点诱变系统生产突变的表达载体。然后,利用“1.4.2”中的表达方法对突变体进行表达和纯化。最后通过SDS-Page和IC-ELISA鉴定并分析ScFv突变体。
1.4.5 建立间接竞争ELISA法。基于纯化后的抗NEO-ScFv建立IC-ELISA检测方法。简而言之,在96孔板上每孔加入包被原NEO-OVA溶液100 μL,放置在4 ℃下过夜;然后用PBST溶液洗涤,在37 ℃下用每孔300 μL的5% PBSM(PBS中5%脱脂奶粉)封闭1 h。用CBS将NEO标准品稀释为一系列标准浓度(200、100、80、50、20、10、5、2、1和0.1 ng/mL),分别与抗NEO-ScFv进行混合后加入孔中(100 μL/孔)。放置在37 ℃下孵育1 h,然后用PBST洗涤。将与辣根过氧化物酶(100 μL /孔;在PBS中稀释1/2 000)缀合的抗生物素蛋白加入孔中,并在37 ℃下孵育30 min。然后用PBST洗涤5次,并添加100 μL /孔的TMB底物,在黑暗中于室溫孵育 10 min 后,加入浓度为2 mol/L的浓硫酸(50 μL /孔)终止显色反应。最后,使用自动酶标仪(Thermo,USA)在450 nm处测量吸光度。以IC 50,测定动态范围和检测下限(LOD)作为评估 IC-ELISA的标准。使用公式(P-S-N)/(P-N)×100%计算抗NEO-ScFv的IC 10、IC 20,IC 50和IC 80的抑制率,其中P是阳性样品的OD 450(50 μL抗NEO ScFv与50 μL CBS混合),S为标准品的OD 450(50 μL ScFv与50 μL NEO标准品的连续浓度混合),N为阴性对照的OD 450(100 μL CBS)。
1.4.6 交叉反应率分析。
配制不同浓度的庆大霉素、卡那霉素和链霉素标准溶液(200.0、100.0、80.0、50.0、20.0、10.0、5.0、2.0、1.0和 0.1 ng/mL)。用确定的最适抗原抗体浓度进行IC-ELISA试验,步骤同“1.4.5”(在加入抗新霉素ScFv时,先将ScFv分别与不同浓度的庆大霉素、卡那霉素和链霉素进行室温孵育 30 min),每个浓度重复3次。然后用SPSS软件计算IC 50,最后根据公式CR=IC 50(新霉素)/IC 50(竞争物)×100%计算交叉反应率。
2 结果与分析
2.1 ScFv基因的构建 VH、VL和ScFv基因的扩增结果如图1所示,分别长约350、330和780 bp。VH基因长度略大于VL基因,与预期结果相似,经过NCBI BLAST对比可知为抗体可变区基因,由此可知,成功扩增和组装ScFv基因。用Sfi Ⅰ和Not Ⅰ消化ScFv基因和pCANTAB5E载体,用T4 DNA连接酶连接产物后,将重组质粒转化到大肠杆菌TG1细胞中进行涂板培养。
2.2 单链抗体的表达和纯化
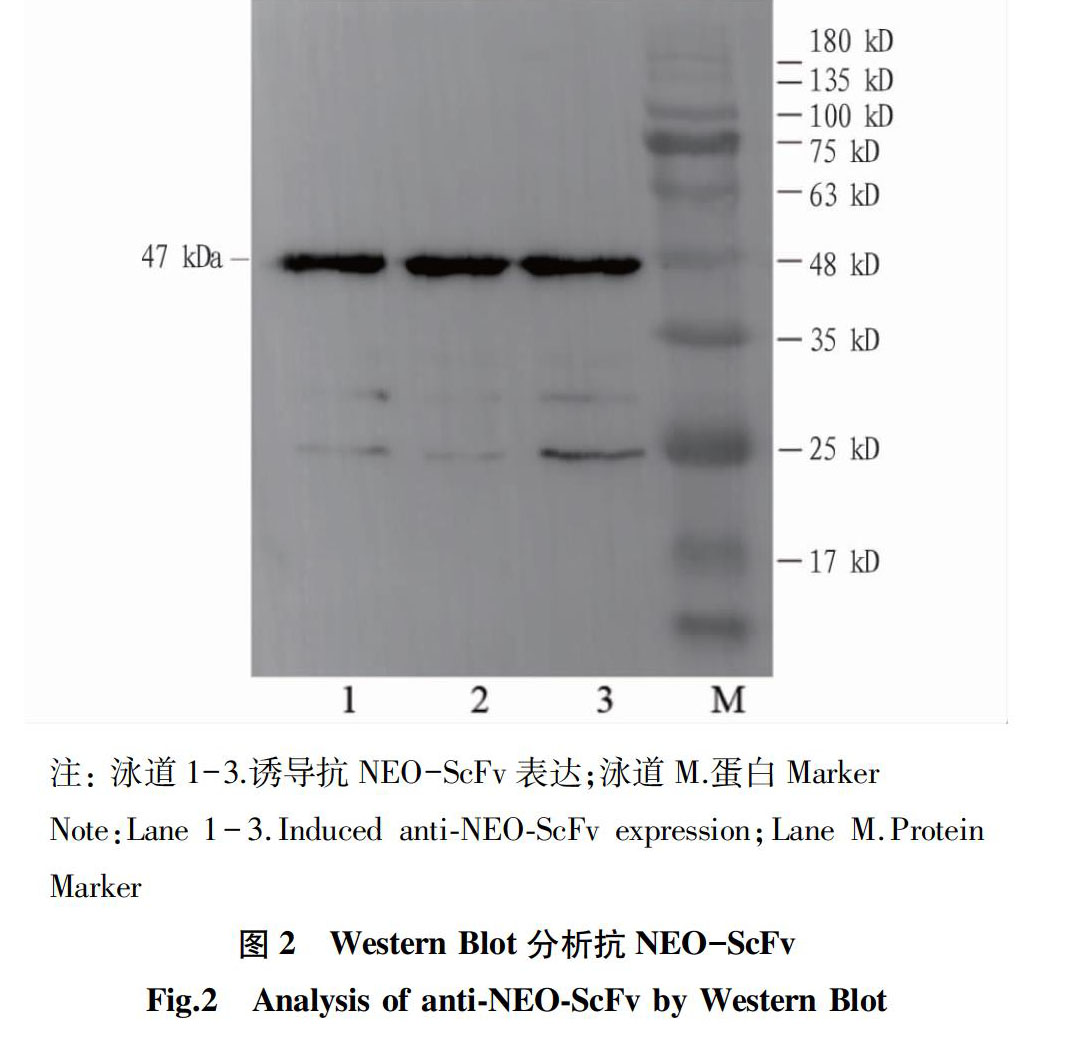
对得到的重组质粒ScFv进行测序和表达,在37 ℃下加入1 mmol/L的IPTG诱导表达,并用Western Blot检测(图2),结果表明ScFv抗体大小为 47 kD,与预测一致,说明成功表达抗NEO-ScFv。然后使用Beaver Beads TM His-tag蛋白纯化磁珠进行纯化(图3),说明ScFv蛋白大部分存在上清液中,分别用20、100、200、 250 mmol/L咪唑洗涤后,发现用20 mmol/L咪唑洗脱时可以将大部分单链抗体洗脱,其上清液即为纯化的单链抗体蛋白。最后将纯化后的蛋白在-20 ℃下保存。
2.3 同源建模和分子对接
通过NCBI Blast序列比对,发现抗NEO-ScFv序列与蛋白质PDB ID:5i4f.1.A有高度序列一致性(67.07%),可作为模板进行同源建模。随后,利用SWISS-MODEL同源性建模软件建立单链抗体模型。然后用Procheck、Verify-3D和Errat对模型合理性进行评价。Ramachandran图结果显示模型中93.3%的氨基酸残基位于核心区域,3.9%位于允许区域,仅有1.7%位于不合理区域内。说明该模型中98.3%蛋白质残基在合理范围内,符合立体化学能定律。Verify-3D的结果表明,ScFv模型中94.3%氨基酸残基的平均3D-1D得分大于0.2,因此该模型通过了Verify-3D测试。Errat结果表明,总体质量因子为89.579。所以,构建的抗NEO-ScFv模型具有很高的可靠性,可用于与NEO抗原的分子对接。
Leu229组成。NEO可以同时与残基Glu51、Glu36、Glu100、Gly224、Cys167、Asn102和Asp183形成相对稳定的氢键网络体系。此外,(2S,3S,4R,4R,5R,6R)-吡喃环上的5-氨基基团还可以与残基Tyr165形成π-π堆积作用。然而,发现疏水性的残基Leu227靠近NEO分子中的(2R,3R,4R,4R,5S,6R)-吡喃环上的亲水性基团6-氨甲基和5-羟基。初步推测,如果将残基Leu227转化为亲水氨基酸,它将促进NEO与ScFv的结合。有趣的是,当残基Leu227突变为Asp227时,总结合能从-21.306降至-32.776 kJ/mol。此时,6-氨甲基基团可以与Asp227形成氢键作用。(2R,3R,4R,4R,5S,6R)-吡喃环上,质子化的3-氨基基团可以与Trp34形成Pi-H相互作用。这些结果表明,突变后的单链抗体与NEO之间的结合更加稳定。因此,在该研究中Leu227被Asp取代,用于制备ScFv突变体。
2.4 建立IC-ELISA检测结果 用IC-ELISA方法建立的标准曲线如图5所示,相关线性回归方程为y=-0.186 5x+ 0.558 0(R2=0.989 9)。IC 50为1.58 ng/mL,最低检测线(IC 10)为 19.84~285.60 ng/mL,相比于未突变前的亲代ScFv的IC 50: 26.23 ng/mL,灵敏度提高了16.6倍。说明经过分子对接和定向进化,获得了灵敏度更高的抗新霉素的单链抗体,并建立了间接竞争ELISA检测方法,为食品中的新霉素残留提供了灵敏度更高的检测方法。
2.5 交叉特异性分析
建立的IC-ELISA检测方法的特异性取决于抗NEO-ScFv的交叉反应性,试验选取与新霉素结构类似的庆大霉素、卡那霉素和链霉素为竞争物,用已建立的新霉素IC-ELISA检测方法检测竞争物,其交叉反应性见表2,结果表明建立的新霉素IC-ELISA检测方法具有良好的特 异性。
3 讨论与结论
目前,针对动物源性食品抗生素残留的检测方法主要为超高效液相色谱分析法,但该方法所需仪器昂贵并且操作复杂,浪费时间和人力,适合对可疑样品进行确证检测;而免疫分析法因为操作简便、节约时间而适合对大批量样品进行初步筛查。免疫法中需要利用的抗体是制约发展的瓶颈之一,随着基因工程技术的不断进步,单链抗体的出现解决了这一问题。单链抗体是仅具有一条链的合成抗体,对抗原具有特定的亲和力。与常规的单抗和多抗相比,单链抗体可以在分子水平上进行研究,并且可以通过基因突变进行改造以提高其抗原结合亲和力 [15]。
该项研究中,在已有能分泌抗新霉素单克隆抗体的杂交瘤细胞中提取了总RNA,然后反转录为cDNA。使用cDNA作为模板扩增VH和VL编码序列,并添加互补的接头序列。通过SOE-PCR拼接整个ScFv基因,然后将基因转入大
肠杆菌表达系统进行表达,得到抗NEO初代的单链抗体。为进一步提高其灵敏度,通过同源建模和分子对接等方式,分析两者结合的主要作用力和抗体与小分子NEO结合的关键氨基酸残基,并进行定向改造,从而得到亲和力更高、灵敏度更强的抗NEO-ScFv突变体。以突变体为基础,建立间接竞争ELISA检测方法,其IC 50达1.58 ng/mL,与初代相比灵敏度提高了16.6倍。该研究为动物性食品在上市前检测新霉素残留提供了灵敏度更高、特异性更强的检测方法。
参考文献
[1]
古丽努尔·阿巴斯.新霉素在兽医方面的应用[J].当代畜牧,2016(29):120.
[2] 张静,赵立军,李云.HPLC-MS/MS法测定饲料中硫酸新霉素的含量[J].饲料研究,2017(11):27-32.
[3] 李爽,张燕,张洁琼.鸡蛋细菌耐药性检测试验报告[J].科学种养,2020(8):45-46.
[4] 高月,王耀,胡骁飞,等.氨基糖苷类药物的危害及其检测方法研究进展[J].河南农业科学,2016,45(6):9-14.
[5] 欧盟:修订新霉素在动物源食品上的残留限量[J].中国标准导报,2013(11):15.
[6] 黄敏文,侯玉荣,张倩,等.高效液相色谱-脉冲安培电化学检测器法替代微生物检定法测定硫酸新霉素效价的研究[J].中国药学杂志,2020,55(2):133-140.
[7] 刘庄蔚,朱健萍,梁秋霞,等.HPLC-CAD法测定硫酸新霉素中新霉素B、新霉素C、新霉胺及其他有关物质[J].中国抗生素杂志,2019,44(3):370-375.
[8] 夏玲玲.高效液相色谱-串联质谱法建立同时测定肌肉组织中4种药物残留的方法及其确认[J].科技资讯,2020,18(3):230,232.
[9] 孙明君,邱芳,张金玲,等.进出口动物中新霉素ELISA检测方法的建立[J].中国畜牧兽医,2016,43(12):3163-3169.
[10] 刘沙洲,桑小雪,欧阳华学,等.新霉素ELISA检测方法的建立[J].食品科学,2011,32(14):227-231.
[11]
王新桐,孙佳芝,高丽丽,等.转基因棉花中新霉素磷酸转移酶(NPTⅡ)双抗体夹心ELISA定量检测方法的建立[J].农业生物技术学报,2014,22(3):372-379.
[12] 刘淑华,何方洋,冯才伟,等.牛奶中新霉素残留胶体金免疫层析快速检测技术的研制[J].食品工业科技,2013,34(6):73-76.
[13] 刘宣兵,滕蔓,张改平,等.新霉素单克隆抗体的制備及其免疫学特性鉴定[J].华北农学报,2009,24(4):80-83.
[14] HE X,DUAN C F,QI Y H,et al.Virtual mutation and directional evolution of anti-amoxicillin ScFv antibody for immunoassay of penicillins in milk[J].Anal Biochem, 2017, 517:9-17.
[15] WANG J P,DONG J,DUAN C F,et al.Production and directional evolution of antisarafloxacin ScFv antibody for immunoassay of fluoroquinolones in milk[J].J Agric Food Chem,2016,64(42):7957-7965.
- 稻鳖共生 1亩收入1.6万元
- 全产业链开发独家核桃年卖800万元
- 中国泡菜王:坚持传统制作方法,年销3000万元
- 两坛特产打天下一坛冬瓜猪,一坛土家酒
- 巧卖百香果 年销过千万元
- 石粉变胶 让发大财的梦想飞起来
- 石粉变塑料 变出财富和快乐
- 盆栽《五朵金花》富万家
- 推广15年的绿色新能源—氢能燃油
- 霹雳闪光豆 沙子卖天价 政府补贴800元
- 安全交易平台
- 想卖货挣钱 要选对产品 用对方法
- 今日解密
- 迷你(Mini)影像馆
- 青岛市亿城科瑞建材制品厂
- “有技无证、拿医师证” 找二十四气医学培训
- 日用品维修
- 无店铺经营月子汤二胎政策让你更赚钱
- 草原天然药材 蒙古特色疗法效果好
- 发家致富好项目 古老工艺铺财路
- 暴利的地方性棋牌
- 马云门徒 卷款私逃
- 创业公司人才管理的经验
- 劳务派遣发生矛盾向谁维权
- 婚礼状况频出婚庆公司赔钱
- framableness's
- frame
- frameableness
- frameablenesses'
- frameablenesses
- frameableness's
- framed
- frameless
- frame of mind
- framer
- framers
- framer's
- frames'
- frames
- framework
- frameworkagreement
- framework aˌgreement
- frameworks
- frame²
- frame¹
- framing
- franc
- franchisabilities
- franchisability
- franchisable
- 阿斗式的人物
- 阿斗当太子——扶不起来
- 阿斗当官——有名无实
- 阿斗当皇帝
- 阿斗有权,诸葛有能
- 阿斗的江山
- 阿斗的江山——白送
- 阿斯匹林
- 阿斯旺
- 阿时
- 阿时趋俗
- 阿昌族
- 阿昌族的传统节日风俗故事
- 阿是穴
- 阿曲
- 阿曼
- 阿替五替
- 阿木林
- 阿本郎
- 阿朮
- 阿权膴仕
- 阿杜
- 阿条
- 阿枉
- 阿柄