美丽异木棉花芽分化期形态结构初步研究
张永强+沈海岑+刘念
摘要:美丽异木棉Ceiba speciosa因其花色鲜艳、花期长而被广泛种植应用于园林绿化。为探析美丽异木棉花芽分化特征以便于指导花期调控相关研究,本文根据花芽大小,将其分为10级,通过应用石蜡切片法,结合花芽不同发育时期的外部形态特征,以期找出花芽发育时期与形态特征之间关系。美丽异木棉花芽生长发育过程中花芽的长宽比呈现先下降1~3级再上升4~10级的特点。其花芽分化期与形态特征对应关系为:前分化期(枝端生长点)、萼片分化期(枝端生长点)、花瓣形成期(1~2级)、雄蕊附属物及子房形成期(3~4级)、雌雄蕊形成期(4级)、胚珠形成期(5~6级)共6个时期。
关键词:美丽异木棉;花芽分化;发育时期;形态结构
中图分类号:S685
文献标志码:A
文章编号:1671-2641(2017)06-0068-05
收稿日期:2017-11-17
修回日期:2017-12-14
Abstract: Ceiba speciosa has great ornamental characteristics in variety color and long florescence, which has been widely applied in landscaping. In order to know the characteristics of flower bud differentiation in Ceiba speciosa, which were to guide the relative research on blooming control. In this paper, according to the dimension of flower buds, divided them into 10 stages. And applied the paraffin method, combined with morphological structures character in different stage, to find out the relationship with the flower buds development stage and morphological structures character. During the development of flower bud, the length and width ratio showed declined in early stage(1st~3rd stage) and ascend in 4th~10th stage. And the characters of flower bud differentiation in six periods were showed as below: the predifferentiation period (growing point on the top of branch); the calyx lobe differentiation period (growing point on the top of branch); the formative period of petals (1st~2nd stage); the appurtenance of stamen period and the formative period of ovary (3rd~4th stage); the formative period of gynoecium and androecium (4th stage) and the formative period of ovule (5th ~ 6thstage).
Key words: Ceiba speciosa; Flower bud differentiation; Development stage; Morphological structure
1 前言
美丽异木棉Ceiba speciosa,又称酒瓶木棉、美丽吉贝、青皮木棉等,是木棉科Bombacaceae吉贝属Ceiba植物,原产于南美洲的阿根廷东北部、玻利维亚东部、巴拉圭、乌拉圭和巴西南部等地[1~2]。广东于20世纪70年代末80年代初开始引种,90年代初广州市园林科学研究所进行了相关研究之后并开始大力繁殖推广[3]。其树形优美,飘逸飒爽,花期较长且开花时间差异較大,盛花时花多叶少,花色艳丽,极为美观,是优良的观赏树木 [4~5]。
所谓花芽分化是指植物生长到一定阶段后,叶芽生理和组织状态转变为花芽生理和组织状态,继而发育成为花器官雏形的过程[6]。花芽分化直接影响到植物的开花时间,人们通常采取控制花芽分化进行花期调控,以达到优产丰产的目的[7]。何见等[8]对光皮树Swida wilsoniana的花芽分化的形态和解剖结构进行了观察和研究,将光皮树花芽分化分为5个时期。贺娜等[9]对油橄榄Olea europaea的花芽分化的形态解剖学特征进行了观测和研究,确定了油橄榄花芽形态分化的各时期特征。花芽分化和发育是开花植物发育中最关键的时期,了解花芽分化机理,掌握花芽分化规律,在花期调控方面有着重要的指导意义。目前国内外对果树的花芽分化研究报道较多[10~13],有关观赏树种的花芽分化研究较少,而关于美丽异木棉花芽形态分化特征的研究尚未见报道。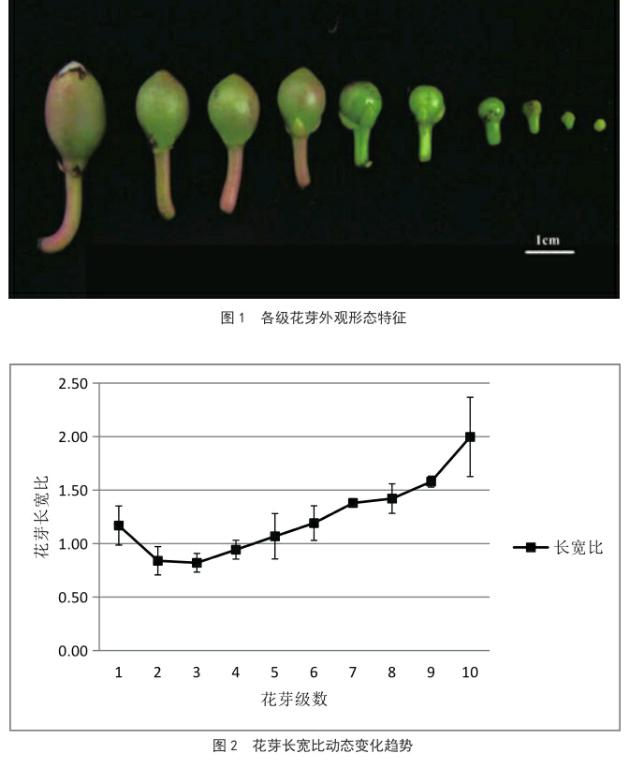
本文以广州市白云区陈田花园内的美丽异木棉为研究对象,研究美丽异木棉花芽分化各个阶段特征及其与外观形态变化的关系,旨在了解和掌握美丽异木棉的花芽分化规律,以便为后期美丽异木棉的花期调控工作提供理论基础和指导。
2材料与方法
2.1试验材料
本研究的试验对象种植于广东省广州市白云区陈田花园(23°12′40″N;113°17′13″E)。各采取3株生长发育正常、长势良好且无病虫害的10个不同大小时期的美丽异木棉花芽,每个时期采取10个样本。
2.2成熟花朵的解剖及花芽形态观察与测定
为了解并判定花芽不同分化时期的结构特征,对美丽异木棉成熟开放的花朵进行解剖观察并拍照记录。应用游标卡尺对不同大小时期的花芽进行长、宽的测定,应用SPSS 19.0的ANOVO单因素分析法,对不同时期花芽的长、宽进行显著差异分析;探究不同大小时期的花芽长宽比的变化规律。
2.3石蜡切片的制作与观察
将采集不同大小时期的花芽及枝端生長点样本于FAA固定液中真空抽滤后固定24 h以上并转入70%酒精保存。由于美丽异木棉花芽外部萼片较厚,本试验的石蜡切片的制法在参照Lai 等[14]的方法基础上将浸蜡方式改为3次,每次5 h;且将5级以上的花芽剥去萼片进行试验处理,以便石蜡浸透样本。切片厚度为8 μm,应用苏木精-伊红染液进行染色,并于用LeicaDM5500B型显微镜观察花芽纵切面的解剖结构并拍摄照片,记录美丽异木棉花芽分化过程中解剖结构的变化特征。
3结果与分析
3.1花芽发育的形态变化特征
参照前人[14~16]关于花芽分化的研究成果,结合美丽以木棉花芽发育特点,将花芽以直径大小为依据划分为1至10级,花芽外观如图1所示。不同级别花芽的大小形态由表1可知,除8、9两级花芽外,其它各级花芽之间的直径(宽)均在p≤0.05水平上存在显著性差异,而花芽的高(长)除1、2、3级外,其它各级之间在p≤0.05水平上存在显著性差异。
花芽大小在1~3级时,外部变化表现为直径增长较快,由1.88 mm增长至3.68 mm;长度的增长较慢,由2.14 mm增至3.01 mm;花芽长宽比呈下降趋势(图2)。至发育后期(8~10级),各级花器官趋于成熟,花芽的直径增长变慢,因此8、9两级花芽的直径不存在显著差异。总体上看,从可见的花芽至开花前,美丽异木棉花芽的平均直径由1.88 mm增长至11.70 mm,高由2.14 mm增长至23.41 mm;1~3级的花芽长宽比表现为下降趋势,在3~10级则呈现出上升趋势。
花芽大小在1~5级的阶段时,花芽外观呈绿色,6级时花芽萼片开始出现红色,随着花芽的发育红色加深、面积增大;1~3级的花芽横向变宽,顶端较平,呈扁球形;4~6级的花芽,其形态纵向生长变快,第4级花芽形状接近圆形;第7级的花芽顶端出现小突起,随着花芽进一步发育,突起变尖;第10级花芽的萼片顶端突起裂开,露出花瓣。
3.2美丽异木棉的花芽分化特征
根据2016年的观测记录,美丽异木棉每年七月底至八月初花芽开始形态分化。应用石蜡切片法对不同时期大小的花芽进行切片观察;同时取美丽异木棉成熟花朵进行解剖(图3)。根据成熟花的结构判断切片中花芽分化的各个时期,美丽异木棉花芽分化特征可划分为6个时期。
1)前分化期(图4-A,枝端生长点):该时期处于花芽分化前期,此时分生组织分裂速度加快,生长点变为圆滑肥大,体积增大,呈半球形。花芽外观形态肉眼不可见,存在于枝端生长点或叶腋中。
2)萼片形成期(图4-B,枝端生长点):形态分化初期,生长点先增大伸长,而后在周围产生突起,向内弯曲,继而生长点顶端变得宽而平坦,萼片继续生长并覆盖生长点。此时花芽外观形态肉眼不可见,存在于枝端生长点或叶腋中。
3)花瓣形成期(图4-C、图4-D,第1~2级花芽):随着萼片的分化和不断发育,在生长点处形成几个小突起,以不同速度向上生长,最靠近花萼的突起为花瓣原基(图4 c),花瓣原基进一步伸长发育为花瓣(图4 d),最终形成五个花瓣,五枚花瓣在花萼的压力下,层层覆盖在中间的生长点上。此时在枝头顶端和叶腋能看到针孔大小的绿色花芽。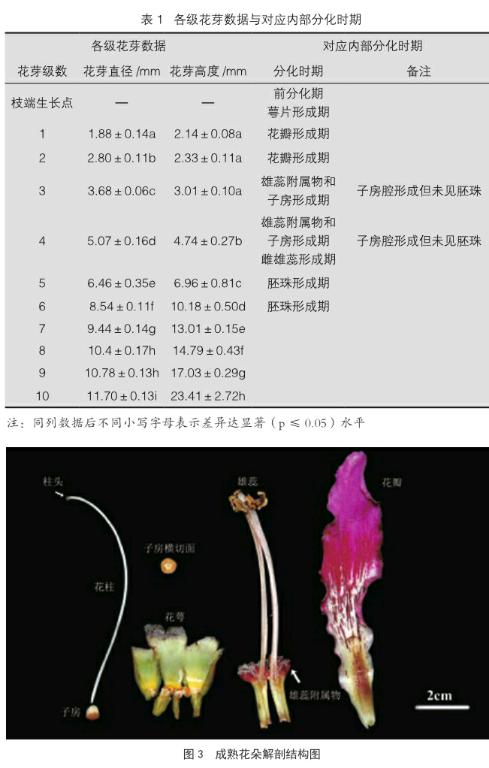
4)雄蕊附属物和子房形成期(图4-E、图4-F,第3~4级花芽):花瓣形成后,生长点继续分化,上部逐渐变尖,出现若干个小突起,这些突起即为雄蕊附属物原基,下部子房壁开始分离,子房腔逐渐形成,但未见成型胚珠。外部形态观察发现,花芽外部为绿色萼片。
5)雌雄蕊形成期(图4-G,第4级花芽):雄蕊附属物原基逐渐发育为成型的雄蕊附属物,随后其中间伸长,雄蕊原基(图4 g)逐渐从雄蕊附属物中间伸长,并在顶端形成小突起,雌蕊原基被包裹其中。此时花芽逐渐伸长,外部形态由近圆形变为椭圆形,外部萼片由深绿色变为浅绿色。
6)胚珠形成期(图4-H,第5~6级花芽):雌雄蕊形成后,雌蕊下部形成5个子房腔,每个子房腔有多个胚珠(图4 h)。此时花芽的高度和直径都继续增加,但花芽高增加速度更快,外部形态表现为花芽继续伸长,顶端突起变尖,在直径最大处的萼片上显现红色。
4结论与讨论
不同植物进入分化期的时间与各分化期对应的内外形态结构存在明显差异[17~18]。通常情况下,花芽分化大致可分为分化初期、萼片分化期、花瓣分化期、雄蕊分化期、雌蕊分化期5个阶段,各个阶段的命名略有差异[19~20]。美丽异木棉整个分化期可分为前分化期(枝端生长点)、萼片分化期(枝端生长点)、花瓣形成期(1~2级)、雄蕊附属物及子房形成期(3~4级)、雌雄蕊形成期(4级)、胚珠形成期(5~6级)6个时期,与外部分级相对应时,存在交错重叠的现象。李永涛等[21]等在对早实核桃Juglans regia花器官发育的研究中,进行了详细的划分,其中包括胚珠分化期。大部分学者的研究都未将胚珠形成期单独划分出来,徐桂香等[22]在对紫杏Armeniaca Dasycarpa花芽分化进行研究时,将其花芽分化的过程分为未分化期、分化初期、萼片分化期、花瓣分化期、雄蕊分化期和雌蕊分化期6个时期,与本文中对美丽异木棉花芽分化时期的划分有所区别,但同样存在各分化时期之间交叉重叠的现象。
不同发育时期的花芽其长宽比变化具不同特征。朱雯等[20]的研究结果表明广宁红花油茶Camellia semiserrata的花芽长宽比随着分化时期的变化由大变小再变大;王湘南等[16]对油茶Camellia oleifera的的研究结果表明花芽在发育过程中,长宽比存在由小变大再减小再增大的过程。本研究中,美丽异木棉的花芽长宽比在发育的过程中呈现出先下降后上升的变化特点。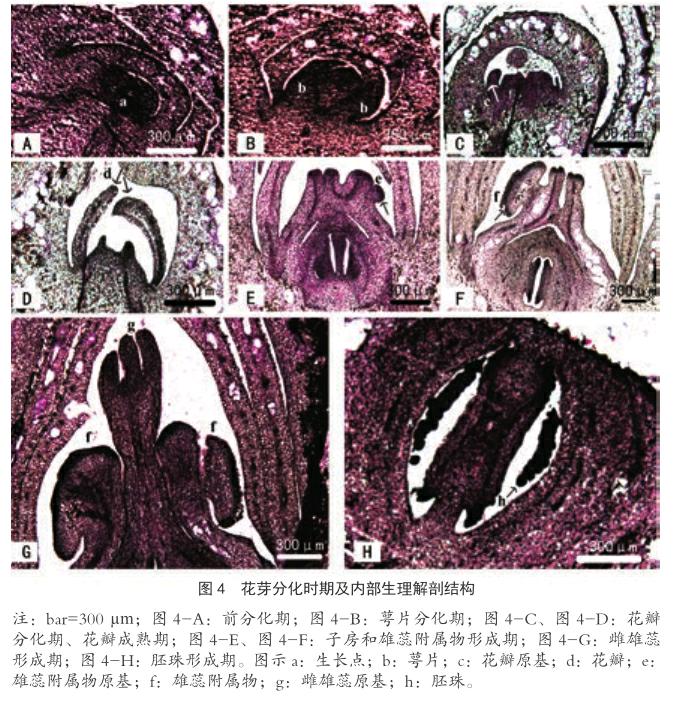
研究发现,不同大小的美丽异木棉花芽的外部形态与不同的分化期存在一定的联系。花芽的外部形态与分化期相对应,前分化期及萼片分化期的花芽位于叶腋和枝端生長点,肉眼不可见;花芽生长到1~2级大小时进入花瓣形成期,花芽达到3~4级时进入雄蕊附属物和子房形成期,雌雄蕊形成期出现在4级花芽中,5~6级花芽出现胚珠。花芽分化至7、8级时基本完成分化,形成花器官雏形。
根据花芽分化的时期与外部形态特征对应的关系,掌握这一规律将有助于从美丽异木棉花芽外部形态特征了解其内部的分化时期,以便及时采取措施进行花期管理或促使其花芽整齐分化,达到使其整齐开花的目的。
参考文献:
[1] Gibbs P.,Semir J. A taxonomic revision of the genus Ceiba Mill.(Bombacaceae)[J]. Anales del Jardín Botánico de Madrid,2003,60(2):259-300.
[2] 张方秋,朱报著,潘文,等. 美丽异木棉化形态结构特征和观赏类型划分[J].广东农业科学,2015(14):48-52.
[3] 李虬,陈惠明. 木棉科植物引种及繁殖研究[J]. 广东园林,1996,18(4):10-13.
[4] 臧小平. 美丽吉贝主要特性、园林绿化应用与播种繁殖[J]. 亚热带农业研究,2005,1(1):34-36.
[5] 肖珍泉,刘湘源,费胜强,等. 美丽异木棉的种植繁殖与栽培管理[J]. 广东园林,2006,28(5):45-46.
[6] 曾辉,杜丽清,邹明宏,等. 澳洲坚果花芽分化期间内源激素的变化[J]. 安徽农业科学,2008,36(34):14949-14953.
[7] 张彦卿,齐国辉,李保国,等. 黄连木雌花芽形态分化研究[J]. 西北植物学报,2011,31(5):972-976.
[8] 何见,蒋丽娟,李昌珠,等. 光皮树花芽分化的形态和解剖特征观察[J]. 植物资源与环境学报,2009,18(2):57-61.
[9] 贺娜,徐田,宁德鲁,等. 油橄榄花芽分化的形态解剖学特征观测[J]. 林业调查规划,2016,41(2):82-85.
[10] 李正理. 植物制片技术[M]. 北京:北京科学出版社,1991:71,86,138,148.
[11] 许明宪,黄尚志. 苹果花芽的生理分化和形态分化[J]. 园艺学报,1962,1(2):137-152.
[12] 蔡兴元,虞锦星,褚孟螈,等. 李花芽分化物候期的观察[J]. 中国果树,1984,2(5):15-16,37.
[13] 赵文东,孙凌俊,徐静,等. 薄膜温室葡萄花芽分化规律的研究[J]. 果树学报,2006,23(1):9-12.
[14] Hang Gui Lai,Xia Chen,Zheng Chen,et al. Induction of female 2n gametes and creation of tetraploids through sexual hybridization in cassava (Manihot esculenta) [J]. Euphytica,2015,201(2):265-273.
[15] 祁宏英,姚美玲,徐洪国. 龙葵花芽分化形态解剖学研究[J]. 北方园艺,2017(9):135-138.
[16] 王湘南,蒋丽娟,陈永忠. 油茶花芽分化的形态解剖学特征观测[J]. 中南林业科技大学学报,2011,31(8):22-27.
[17] Tomasz L. Woznicki,Ola M. Heide,Siv F. Remberg,et al. Effects of controlled nutrient feeding and different temperatures during floral initiation on yield,berry size and drupelet numbers in red raspberry ( Rubus idaeus L.) [J]. Scientia Horticulturae,2016,212:148-154.
[18] 袁德义,邹锋,谭晓风,等. 油茶花芽分化及雌雄配子体发育的研究[J].中南林业科技大学学报,2011,31(3):65-70.
[19] 郝敬虹,齐红岩,阎妮,等. 园艺作物花芽分化的研究进展[J]. 农业科技与装备,2008,29(1):7-9.
[20] 朱雯,许逸林,李文锋,等. 广宁红花油茶花芽分化特征[J]. 广东农业科学,2016,51(3):51-54.
[21] 李永涛,赵勇刚,杨克强,等. 早实核桃花器官发育的解剖学研究[J]. 园艺学报,2011,38(3):434-440.
[22] 徐桂香,廖康,赵世荣,等. 紫杏花芽形态分化特性研究[J]. 新疆农业科学,2016,53(4):604-609.
摘要:美丽异木棉Ceiba speciosa因其花色鲜艳、花期长而被广泛种植应用于园林绿化。为探析美丽异木棉花芽分化特征以便于指导花期调控相关研究,本文根据花芽大小,将其分为10级,通过应用石蜡切片法,结合花芽不同发育时期的外部形态特征,以期找出花芽发育时期与形态特征之间关系。美丽异木棉花芽生长发育过程中花芽的长宽比呈现先下降1~3级再上升4~10级的特点。其花芽分化期与形态特征对应关系为:前分化期(枝端生长点)、萼片分化期(枝端生长点)、花瓣形成期(1~2级)、雄蕊附属物及子房形成期(3~4级)、雌雄蕊形成期(4级)、胚珠形成期(5~6级)共6个时期。
关键词:美丽异木棉;花芽分化;发育时期;形态结构
中图分类号:S685
文献标志码:A
文章编号:1671-2641(2017)06-0068-05
收稿日期:2017-11-17
修回日期:2017-12-14
Abstract: Ceiba speciosa has great ornamental characteristics in variety color and long florescence, which has been widely applied in landscaping. In order to know the characteristics of flower bud differentiation in Ceiba speciosa, which were to guide the relative research on blooming control. In this paper, according to the dimension of flower buds, divided them into 10 stages. And applied the paraffin method, combined with morphological structures character in different stage, to find out the relationship with the flower buds development stage and morphological structures character. During the development of flower bud, the length and width ratio showed declined in early stage(1st~3rd stage) and ascend in 4th~10th stage. And the characters of flower bud differentiation in six periods were showed as below: the predifferentiation period (growing point on the top of branch); the calyx lobe differentiation period (growing point on the top of branch); the formative period of petals (1st~2nd stage); the appurtenance of stamen period and the formative period of ovary (3rd~4th stage); the formative period of gynoecium and androecium (4th stage) and the formative period of ovule (5th ~ 6thstage).
Key words: Ceiba speciosa; Flower bud differentiation; Development stage; Morphological structure
1 前言
美丽异木棉Ceiba speciosa,又称酒瓶木棉、美丽吉贝、青皮木棉等,是木棉科Bombacaceae吉贝属Ceiba植物,原产于南美洲的阿根廷东北部、玻利维亚东部、巴拉圭、乌拉圭和巴西南部等地[1~2]。广东于20世纪70年代末80年代初开始引种,90年代初广州市园林科学研究所进行了相关研究之后并开始大力繁殖推广[3]。其树形优美,飘逸飒爽,花期较长且开花时间差异較大,盛花时花多叶少,花色艳丽,极为美观,是优良的观赏树木 [4~5]。
所谓花芽分化是指植物生长到一定阶段后,叶芽生理和组织状态转变为花芽生理和组织状态,继而发育成为花器官雏形的过程[6]。花芽分化直接影响到植物的开花时间,人们通常采取控制花芽分化进行花期调控,以达到优产丰产的目的[7]。何见等[8]对光皮树Swida wilsoniana的花芽分化的形态和解剖结构进行了观察和研究,将光皮树花芽分化分为5个时期。贺娜等[9]对油橄榄Olea europaea的花芽分化的形态解剖学特征进行了观测和研究,确定了油橄榄花芽形态分化的各时期特征。花芽分化和发育是开花植物发育中最关键的时期,了解花芽分化机理,掌握花芽分化规律,在花期调控方面有着重要的指导意义。目前国内外对果树的花芽分化研究报道较多[10~13],有关观赏树种的花芽分化研究较少,而关于美丽异木棉花芽形态分化特征的研究尚未见报道。
本文以广州市白云区陈田花园内的美丽异木棉为研究对象,研究美丽异木棉花芽分化各个阶段特征及其与外观形态变化的关系,旨在了解和掌握美丽异木棉的花芽分化规律,以便为后期美丽异木棉的花期调控工作提供理论基础和指导。
2材料与方法
2.1试验材料
本研究的试验对象种植于广东省广州市白云区陈田花园(23°12′40″N;113°17′13″E)。各采取3株生长发育正常、长势良好且无病虫害的10个不同大小时期的美丽异木棉花芽,每个时期采取10个样本。
2.2成熟花朵的解剖及花芽形态观察与测定
为了解并判定花芽不同分化时期的结构特征,对美丽异木棉成熟开放的花朵进行解剖观察并拍照记录。应用游标卡尺对不同大小时期的花芽进行长、宽的测定,应用SPSS 19.0的ANOVO单因素分析法,对不同时期花芽的长、宽进行显著差异分析;探究不同大小时期的花芽长宽比的变化规律。
2.3石蜡切片的制作与观察
将采集不同大小时期的花芽及枝端生長点样本于FAA固定液中真空抽滤后固定24 h以上并转入70%酒精保存。由于美丽异木棉花芽外部萼片较厚,本试验的石蜡切片的制法在参照Lai 等[14]的方法基础上将浸蜡方式改为3次,每次5 h;且将5级以上的花芽剥去萼片进行试验处理,以便石蜡浸透样本。切片厚度为8 μm,应用苏木精-伊红染液进行染色,并于用LeicaDM5500B型显微镜观察花芽纵切面的解剖结构并拍摄照片,记录美丽异木棉花芽分化过程中解剖结构的变化特征。
3结果与分析
3.1花芽发育的形态变化特征
参照前人[14~16]关于花芽分化的研究成果,结合美丽以木棉花芽发育特点,将花芽以直径大小为依据划分为1至10级,花芽外观如图1所示。不同级别花芽的大小形态由表1可知,除8、9两级花芽外,其它各级花芽之间的直径(宽)均在p≤0.05水平上存在显著性差异,而花芽的高(长)除1、2、3级外,其它各级之间在p≤0.05水平上存在显著性差异。
花芽大小在1~3级时,外部变化表现为直径增长较快,由1.88 mm增长至3.68 mm;长度的增长较慢,由2.14 mm增至3.01 mm;花芽长宽比呈下降趋势(图2)。至发育后期(8~10级),各级花器官趋于成熟,花芽的直径增长变慢,因此8、9两级花芽的直径不存在显著差异。总体上看,从可见的花芽至开花前,美丽异木棉花芽的平均直径由1.88 mm增长至11.70 mm,高由2.14 mm增长至23.41 mm;1~3级的花芽长宽比表现为下降趋势,在3~10级则呈现出上升趋势。
花芽大小在1~5级的阶段时,花芽外观呈绿色,6级时花芽萼片开始出现红色,随着花芽的发育红色加深、面积增大;1~3级的花芽横向变宽,顶端较平,呈扁球形;4~6级的花芽,其形态纵向生长变快,第4级花芽形状接近圆形;第7级的花芽顶端出现小突起,随着花芽进一步发育,突起变尖;第10级花芽的萼片顶端突起裂开,露出花瓣。
3.2美丽异木棉的花芽分化特征
根据2016年的观测记录,美丽异木棉每年七月底至八月初花芽开始形态分化。应用石蜡切片法对不同时期大小的花芽进行切片观察;同时取美丽异木棉成熟花朵进行解剖(图3)。根据成熟花的结构判断切片中花芽分化的各个时期,美丽异木棉花芽分化特征可划分为6个时期。
1)前分化期(图4-A,枝端生长点):该时期处于花芽分化前期,此时分生组织分裂速度加快,生长点变为圆滑肥大,体积增大,呈半球形。花芽外观形态肉眼不可见,存在于枝端生长点或叶腋中。
2)萼片形成期(图4-B,枝端生长点):形态分化初期,生长点先增大伸长,而后在周围产生突起,向内弯曲,继而生长点顶端变得宽而平坦,萼片继续生长并覆盖生长点。此时花芽外观形态肉眼不可见,存在于枝端生长点或叶腋中。
3)花瓣形成期(图4-C、图4-D,第1~2级花芽):随着萼片的分化和不断发育,在生长点处形成几个小突起,以不同速度向上生长,最靠近花萼的突起为花瓣原基(图4 c),花瓣原基进一步伸长发育为花瓣(图4 d),最终形成五个花瓣,五枚花瓣在花萼的压力下,层层覆盖在中间的生长点上。此时在枝头顶端和叶腋能看到针孔大小的绿色花芽。
4)雄蕊附属物和子房形成期(图4-E、图4-F,第3~4级花芽):花瓣形成后,生长点继续分化,上部逐渐变尖,出现若干个小突起,这些突起即为雄蕊附属物原基,下部子房壁开始分离,子房腔逐渐形成,但未见成型胚珠。外部形态观察发现,花芽外部为绿色萼片。
5)雌雄蕊形成期(图4-G,第4级花芽):雄蕊附属物原基逐渐发育为成型的雄蕊附属物,随后其中间伸长,雄蕊原基(图4 g)逐渐从雄蕊附属物中间伸长,并在顶端形成小突起,雌蕊原基被包裹其中。此时花芽逐渐伸长,外部形态由近圆形变为椭圆形,外部萼片由深绿色变为浅绿色。
6)胚珠形成期(图4-H,第5~6级花芽):雌雄蕊形成后,雌蕊下部形成5个子房腔,每个子房腔有多个胚珠(图4 h)。此时花芽的高度和直径都继续增加,但花芽高增加速度更快,外部形态表现为花芽继续伸长,顶端突起变尖,在直径最大处的萼片上显现红色。
4结论与讨论
不同植物进入分化期的时间与各分化期对应的内外形态结构存在明显差异[17~18]。通常情况下,花芽分化大致可分为分化初期、萼片分化期、花瓣分化期、雄蕊分化期、雌蕊分化期5个阶段,各个阶段的命名略有差异[19~20]。美丽异木棉整个分化期可分为前分化期(枝端生长点)、萼片分化期(枝端生长点)、花瓣形成期(1~2级)、雄蕊附属物及子房形成期(3~4级)、雌雄蕊形成期(4级)、胚珠形成期(5~6级)6个时期,与外部分级相对应时,存在交错重叠的现象。李永涛等[21]等在对早实核桃Juglans regia花器官发育的研究中,进行了详细的划分,其中包括胚珠分化期。大部分学者的研究都未将胚珠形成期单独划分出来,徐桂香等[22]在对紫杏Armeniaca Dasycarpa花芽分化进行研究时,将其花芽分化的过程分为未分化期、分化初期、萼片分化期、花瓣分化期、雄蕊分化期和雌蕊分化期6个时期,与本文中对美丽异木棉花芽分化时期的划分有所区别,但同样存在各分化时期之间交叉重叠的现象。
不同发育时期的花芽其长宽比变化具不同特征。朱雯等[20]的研究结果表明广宁红花油茶Camellia semiserrata的花芽长宽比随着分化时期的变化由大变小再变大;王湘南等[16]对油茶Camellia oleifera的的研究结果表明花芽在发育过程中,长宽比存在由小变大再减小再增大的过程。本研究中,美丽异木棉的花芽长宽比在发育的过程中呈现出先下降后上升的变化特点。
研究发现,不同大小的美丽异木棉花芽的外部形态与不同的分化期存在一定的联系。花芽的外部形态与分化期相对应,前分化期及萼片分化期的花芽位于叶腋和枝端生長点,肉眼不可见;花芽生长到1~2级大小时进入花瓣形成期,花芽达到3~4级时进入雄蕊附属物和子房形成期,雌雄蕊形成期出现在4级花芽中,5~6级花芽出现胚珠。花芽分化至7、8级时基本完成分化,形成花器官雏形。
根据花芽分化的时期与外部形态特征对应的关系,掌握这一规律将有助于从美丽异木棉花芽外部形态特征了解其内部的分化时期,以便及时采取措施进行花期管理或促使其花芽整齐分化,达到使其整齐开花的目的。
参考文献:
[1] Gibbs P.,Semir J. A taxonomic revision of the genus Ceiba Mill.(Bombacaceae)[J]. Anales del Jardín Botánico de Madrid,2003,60(2):259-300.
[2] 张方秋,朱报著,潘文,等. 美丽异木棉化形态结构特征和观赏类型划分[J].广东农业科学,2015(14):48-52.
[3] 李虬,陈惠明. 木棉科植物引种及繁殖研究[J]. 广东园林,1996,18(4):10-13.
[4] 臧小平. 美丽吉贝主要特性、园林绿化应用与播种繁殖[J]. 亚热带农业研究,2005,1(1):34-36.
[5] 肖珍泉,刘湘源,费胜强,等. 美丽异木棉的种植繁殖与栽培管理[J]. 广东园林,2006,28(5):45-46.
[6] 曾辉,杜丽清,邹明宏,等. 澳洲坚果花芽分化期间内源激素的变化[J]. 安徽农业科学,2008,36(34):14949-14953.
[7] 张彦卿,齐国辉,李保国,等. 黄连木雌花芽形态分化研究[J]. 西北植物学报,2011,31(5):972-976.
[8] 何见,蒋丽娟,李昌珠,等. 光皮树花芽分化的形态和解剖特征观察[J]. 植物资源与环境学报,2009,18(2):57-61.
[9] 贺娜,徐田,宁德鲁,等. 油橄榄花芽分化的形态解剖学特征观测[J]. 林业调查规划,2016,41(2):82-85.
[10] 李正理. 植物制片技术[M]. 北京:北京科学出版社,1991:71,86,138,148.
[11] 许明宪,黄尚志. 苹果花芽的生理分化和形态分化[J]. 园艺学报,1962,1(2):137-152.
[12] 蔡兴元,虞锦星,褚孟螈,等. 李花芽分化物候期的观察[J]. 中国果树,1984,2(5):15-16,37.
[13] 赵文东,孙凌俊,徐静,等. 薄膜温室葡萄花芽分化规律的研究[J]. 果树学报,2006,23(1):9-12.
[14] Hang Gui Lai,Xia Chen,Zheng Chen,et al. Induction of female 2n gametes and creation of tetraploids through sexual hybridization in cassava (Manihot esculenta) [J]. Euphytica,2015,201(2):265-273.
[15] 祁宏英,姚美玲,徐洪国. 龙葵花芽分化形态解剖学研究[J]. 北方园艺,2017(9):135-138.
[16] 王湘南,蒋丽娟,陈永忠. 油茶花芽分化的形态解剖学特征观测[J]. 中南林业科技大学学报,2011,31(8):22-27.
[17] Tomasz L. Woznicki,Ola M. Heide,Siv F. Remberg,et al. Effects of controlled nutrient feeding and different temperatures during floral initiation on yield,berry size and drupelet numbers in red raspberry ( Rubus idaeus L.) [J]. Scientia Horticulturae,2016,212:148-154.
[18] 袁德义,邹锋,谭晓风,等. 油茶花芽分化及雌雄配子体发育的研究[J].中南林业科技大学学报,2011,31(3):65-70.
[19] 郝敬虹,齐红岩,阎妮,等. 园艺作物花芽分化的研究进展[J]. 农业科技与装备,2008,29(1):7-9.
[20] 朱雯,许逸林,李文锋,等. 广宁红花油茶花芽分化特征[J]. 广东农业科学,2016,51(3):51-54.
[21] 李永涛,赵勇刚,杨克强,等. 早实核桃花器官发育的解剖学研究[J]. 园艺学报,2011,38(3):434-440.
[22] 徐桂香,廖康,赵世荣,等. 紫杏花芽形态分化特性研究[J]. 新疆农业科学,2016,53(4):604-609.
