碳茵调控对虾、蟹混养系统微生物群落功能多样性的影响
李杰 田相利 董双林 张凯 王文君 宋娇娇 张庆起

摘要:利用Biolog生态板方法和冗余分析(RDA)等方法,分析了菌碳调控对虾、蟹混养系统水体细菌群落功能多样性的影响,探讨了水体微生物群落多样性与不同环境因子间的关系。实验通过添加益生菌制剂和糖蜜调节养殖水体中菌群结构,共设6个处理,分别是为短小芽孢杆菌(C2)、蜡样芽孢杆菌(C3)、硝化细菌制剂(C4)、EM制剂(C5),以未添加菌制剂的处理为对照组(C1),另外以未添加碳源和菌制剂的处理为对照组(C0)。研究结果表明,不同菌碳下细菌群落对底物碳的利用数量和代谢强度存在较大差异。随着养殖时间的延长,处理组C3的碳代谢活性和对碳源的利用强度总体上高于其它处理组,而处理组C4较低和对照组CO相差不大。PCA分析结果表明,不同处理的细菌群落处于动态变化之中,反映出细菌群落的组成和代谢特征出现了显著差异,其中处理组C3和C4间差异最为显著。处理组C3的多样性指数如Simpson指数Shannon指数和S--E均匀度均较高,而处理组C4则较低。RAD分析表明,碳菌调控条件下,影响细菌群落碳代谢功能的主要环境因子7月为:Chl a、PO4--P、NO2-N、NH4+-N;8月为:NO3--N、NO2-N、TP、TN和PO4-一P;9月为:TP、Chl a、PO4-一P和NO2--N。综合水质指标和细菌群落对碳源的代谢功能的分析结果,本实验初步表明在C/N为15的虾、蟹混养水体中添加蜡样芽孢杆菌,细菌群落的多样性指数及对碳源的代谢功能为最佳。
关键词:碳氮比;细菌群落;BIOLOG;多样性指数;RDA分析
虾、蟹混养是一种重要的人工养殖模式,该模式具有充分利用养殖系统时间和空间资源、降低饵料系数和预防相关动物疾病的发生等优点,能够提供较高的养殖产量和经济效益。人工养殖系统中,生物链短,营养级少,水体中通常会残留大量的残饵、粪便和生物残体等,从而导致氨氮、亚硝氮、硫化氢、胺类等有害物质积累增加,当其超过养殖系统自净能力时,会进一步导致生态环境的恶化以及养殖生物疾病的暴发。
微生物在养殖生态系统的物质循环和能量流动以及净化水环境中具有重要作用。异养细菌可以分解系统中的有机物,加速氮循环,从而降低养殖水体中的碳氮比。然而异养细菌的生长繁殖与水体中碳和氮含量密切相关,根据生物絮团原理,改变水体C/N比可以促进其生长繁殖,从而降低水体氨氮浓度进而对稳定养殖环境发挥调控作用。据报道,施加微生态制剂可对弧菌等致病菌产生拮抗作用,从而有利于芽孢杆菌和硝化细菌等益生菌对养殖水体净化作用及对养殖系统群落结构改善作用的发挥,进而促进养殖动物的健康生长。李卓佳等在虾池中添加芽孢杆菌,降低养殖水体中氨氮、亚硝氮浓度,稳定了养殖环境。张晓阳等通过菌碳调控,降低养殖系统中氨氮、亚硝氮浓度和饵料系数,提高了凡纳滨对虾的成活率及产量。
Biolog-Eco板技术可以反映环境微生物群落特征,主要是根据细菌群落对不同碳源利用程度的差异来表征其功能多样性,现已被广泛用于微生物群落功能多样性的研究。鉴于异样细菌对水体环境的净化作用及弧菌致病作用,本实验采用传统的培养方法分析了碳氮比调控对虾蟹混养系统水体中异养细菌和弧菌数量的变动,利用Biolog-Eco微平板法探讨了不同处理水体中细菌群落功能多样性的变化,并结合RDA分析研究了水体细菌群落碳代谢功能与环境因子之间的关系,以期在微生物群落功能方面为虾、蟹混养系统中应用菌碳调控技术提供一定的参考。
1.材料与方法
1.1实验池塘与围隔
本研究于2014年7-10月在江苏省赣榆县佳信水产开发有限公司海水养殖池塘中进行。实验利用海水池塘陆基围隔进行,面积5m×5m,水深1.2~1.5m,底质为泥沙质。实验围隔结构参见文献。实验期间,养殖水体利用充气泵进行连续充气。
1.2实验设计与管理
1.2.1实验设计实验设置6个处理组,每组设3个平行。实验通过添加菌制剂和糖蜜调节养殖系统,分别命名为C2(短小芽孢杆菌,1×104cfu/mL)、C3(蜡样芽孢杆菌,1×104cfu/mL)、C4(硝化细菌,2×104cfu/mL)、C5(EM菌,1×104cfu/mL),以未泼洒菌制剂的处理为对照组C1,另外设未添加糖蜜和菌制剂的处理为对照组CO。其中,所用短小芽孢杆菌和蜡样芽孢杆菌均由中国海洋大学养殖生态学实验室分离于虾蟹养殖池塘;硝化细菌为德国某品牌氨硝净;EM菌来源于日本某品牌EM菌。
实验围隔中均放养三疣梭子蟹密度为6ind/m2,放养凡纳滨对虾密度为45ind/m2。实验用三疣梭子蟹苗及凡纳滨对虾皆购自连云港赣榆佳信水产开发有限公司。三疣梭子蟹于7月15日放苗,放苗时平均甲宽(7.27±1.91)cm,平均甲长(3.50±0.29)cm,平均体重(19.80±3.27)g;凡纳滨对虾经暂养后于7月20日放苗,放苗时体长(1.37±0.20)cm,平均体重(0.05±0.01)g。于10月5日收获完毕。
根据Avnimelech与卢炳国等总结的生物絮团养殖系统的C/N公式,根据饵料投喂量调整糖蜜的添加量,通过添加糖蜜调控C/N水平为15。三疣梭子蟹饵料选用鲜活蓝蛤(壳肉比为3.27,含水率为78.29%,含碳量32.78%,含氮量9.76%);凡纳滨对虾选用连云港正大农牧有限公司生产的对虾配合饲料(含水率7.00%,含碳量34.65%,含氮量6.72%)。添加碳源为购于山东寿光某公司的糖蜜(总有机碳含量28.91%)。
1.2.2实验管理鲜活蓝蛤每日18:00-19:00投喂1次,投喂量参照周演根等。对虾饵料每天7:00-8:00与18:00-19:00投喂2次。每7天检查生长,根据虾蟹生长情况调整饵料投喂量。
养殖期间围隔不换水,仅补充因蒸发与渗漏造成的损失,水深保持在1.2~1.4 m之间。7月底开始使用增氧机充氧。每晚10:00至次日清晨6:00充气,晴天下午2:00到4:00充气,并根据天气状况调整。随着饵料投喂量与糖蜜添加量的增加,生物絮团逐步形成后调整成全天24 h连续充氧。糖蜜根据投饵量随时调整,充分溶解后泼洒,每2天泼洒一次,天气状况不好不泼洒。
本实验所用蜡样芽孢杆菌、短小芽孢杆菌和硝化细菌活化后,每7~10d泼洒一次,选择在有风的晴天上午,泼洒糖蜜后施用。从2014年7月25日到9月30日,菌剂共泼洒7次。
1.3样品采集与预处理
本实验于2014年7-9月开展。每月下旬,用采水器在实验围隔的3个固定点采集中层水样,混合每个处理的3个重复水样,然后经800目筛绢过滤,取适量混匀水样置于无菌的聚乙烯瓶中,将采集水样于4℃保温箱迅速带回实验室待分析。
1.4日常水质指标测定
每天上午6:00-7:00用温度计测定水温(T),用便携式溶氧仪(SX716,上海三信仪表厂)测定溶氧(DO),每7天用便携式pH计(pH-033,ATC)测pH,手持式折光仪(Reef Octopus)测盐度(S)。
按照《海洋监测规范》(GB/T12763.4-2007)中推荐的方法测定的水体中总氨氮(NH4+-N)、亚硝酸盐(NO-一N)、硝酸盐(NOa—N)、总氮(TN)、活性磷(PO4-P)、总磷(TP)。NH4+-N测定采用靛酚蓝法,NO2-N测定采用重氮一偶氮法,NO2-N测定使用锌镉还原法,PO4-P测定采用钼蓝法,TN和TP采用过硫酸钾(K2S2O8)氧化法。叶绿素a采用金相灿等的方法测定。
1.5Biolog-ECO板反应
Biolog-ECO板含6类碳源共31种底物碳(3个重复)的96孔板。板上含有三个对照孔,根据微生物对碳源底物的利用情况来分析微生物群落碳代谢特征(功能多样性)。本实验将150μL的样品原液或者经无菌0.85 9/6NaCl溶液进行10倍稀释的水样加人ECO板的孔内,避光条件下恒温(28℃)培养,酶标仪分别测点样后7d(0、24、48、72、96、120、144、168h)内590 nm和750nm下的光密度值。
1.6数据处理与统计分析
平均颜色变化率(average well color devel-opment,AWCD)主要用于检测微生物对单一碳底物的利用能力计算方法l_1。]如下:
AWCD=∑[(C1—R)—(Ci—R)750]/n
式中:Ci为i孔碳底物光密度值;R为对照孔碳底物光密度值;n为培养基碳源种类数(取值随碳源种类数的变化而变化)。每个单孔的实际光密度值为该碳底物在590nm与750nm的光密度差值,其中数值取值>0.06。
本研究用培养72h的测定的Biolog数据来计算细菌群落对六大碳源的利用强度、主成分分析及功能多样性指数。其中多样性指数主要包括:Shannon指数(评估细菌的丰度和均度),McIntosh指数(评估细菌群落多维空间多样性),Simpson指数(评估最常见种优势度),S—E均匀度指数(评估细菌均度)。
本研究利用PCA分析和单因子方差分析(One-way ANOVA)来探讨不同处理在相同时间内细菌群落碳代谢特征的差异性。利用Ex-cel2013、SigmaPlotl2.5、SPSSl7.0软件进行数据处理和分析,以P 2.结果 2.1碳氦比调控对虾、蟹混养系统水体中环境因子指标 表1结果显示,除T、pH和S外,不同处理组水体环境因子指标均存在显著差异,但不同指标趋势有所不同。总体上,各菌碳添加组N、P指标含量均显著低于对照组(P<0.05)。其中,NOz—N以C2和C5最低,而除了C2,其它处理组NH4+一N浓度均显著低于对照组;各添加菌碳处理组相比,C1和C2中Chl a和TP显著高于其余处理组(P<0.05)。 2.2碳氮比调控对虾、蟹混养系统水体中细菌群落功能多样性 2.2.1 平均颜色变化率(AWCD) 图1为不同养殖模式水体中细菌群落AWCD值随养殖实验进行的动态变化。在同一时期,不同处理水体中细菌群落利用Biolog板上的碳源总量随着培养时间的进行总体上呈现上升的变化趋势,在不同时期,同一处理细菌群落对碳源总量的利用速度和能力均出现差异。7月,对照组CO的AWCD值最高,显著高于处理组C1和C2(P<0.05),与其余处理组差异不显著(P>0.05)。8月,处理组C3的AWCD值最高,明显高于其它处理组,C2组次之,而对照组CO则降到最低值。9月,处理组C1、C3和C5的AWCD相差不大,水平较高,均显著高于其他处理组(P<0.05)。 2.2.2 对六大碳源的利用变化 图2为不同养殖模式水体细菌对ECO板上不同类型碳源(聚合物、糖、羧酸、酚、胺和氨基酸)的利用情况。根据AWCD值可以看出,水体细菌对碳源利用具有选择性,主要为聚合物、糖类和氨基酸,且在养殖中后期差异显著。7月,以利用聚合物、糖类和氨基酸为主,不同处理对同一类型碳源的利用强度差异不显著(P>0.05)。8月,不同处理对六大碳源的利用强度均显著差异(P<0.05),均主要利用聚合物,糖类和氨基酸,其中处理组C3对这三类碳源的利用强度最高,显著高于对照组CO和C1。9月,不同处理对六大碳源的利用强度差异显著(P<0.05),均以聚合物和糖类的利用为主,其中处理组C1、C3和C5对这两类碳源的利用强度较高,显著高于处理组C4和CO(P<0.05)。 2.2.3对不同碳源利用的主成分分析(PCA分析)
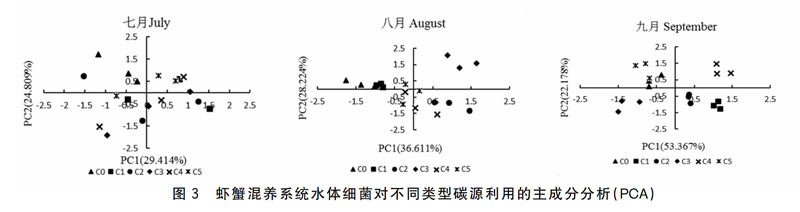
图3为不同养殖模式水体细菌对六大碳源利用的主成分分析结果。7月,六种碳源中提取的两个主成分因子,分别解释变量方差的29.41%和24.81%,其中PC2的得分系数差异显著(P<0.05),表现在对照组CO与处理组C3之间。8月,PCI和PC2分别解释36.61%和28.22%的变量方差,PCI和PC2的得分系数均差异显著(P<0.05),其中PCI得分系数差异表现在:处理组C2和C3与对照组CO和C1之间,PC2表现在:处理组C3与其余处理组之间。9月,PCI和PC2分别解释变量方差为53.37%和22.18%,PCI和PC2的得分系数均差异显著(P<0.05),PCI得分系数差异表现在:各处理间均差异显著(P<0.05),PC2表现在:处理组CO、C4和C5与其余处理组间。
与主成分PCI、PC2具有较高相关系数的碳源(r>0.06)见表2-7月,胺、糖类和氨基酸对PCI起分异作用,表明不同处理水体细菌群落对底物碳的利用在PCI上的显著差异是由这三类碳源导致的。对PC2起分异作用的碳源类型为:聚合物和羧酸。8月,对PCI起分异作用的主要碳源类型为:聚合物、糖类和氨基酸,而对PC2起分异作用的主要碳源类型为:羧酸和胺。9月,聚合物、糖类、羧酸和胺这四类碳源对PCI起分异作用,而酚和氨基酸对这两类碳源PC2起分异作用。
2.2.4利用碳源的多样性指数变化
表3为不同养殖时间各养殖模式水体细菌在培养72 h后利用碳源的多样性指数。结果显示,不同处理水体细菌群落的多样性指数在不同养殖时间呈现不同程度的差异。7月,不同处理水体细菌群落各类多样性指数均无显著差异(P>0.05)。8月,McIntosh指数在各处理间差异显著(P<0.05),且以处理组C3为最高,显著高于其它处理组(P<0.05),而对于Simpson指数、Shannon指数和S-E均匀度指数,各处理间均无显著差异(P>0.05)。9月,各多样性指数在不同处理间均差异显著(P<0.05),其中处理组C3的各多样性指数均为最高,显著高于处理组CO和C4(P<0.05),而Simpson指数、Shannon指数和S-E均匀度指数与处理组C1差异不显著。
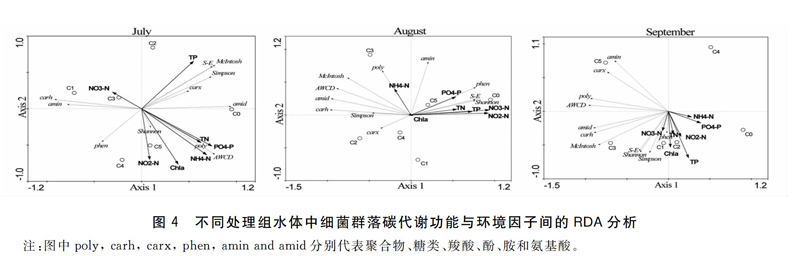
2.3碳氦比调控下虾、蟹混养系统水体中细菌群落的碳代谢功能与环境因子之间的RDA分析
图4为不同模式水体细菌群落的碳代谢功能的时间变化与10种不同环境因子之间的关系。结果显示,随着养殖时间的延长,不同种类的环境因子影响了水体细菌群落的碳代谢功能,且代表不同处理的点分布在排序图的不同空间,说明不同处理组细菌群落存在不同程度的差异。7月,第一排序轴的特征值为53.1%,第二排序轴的特征值为21%,共能解释74.1%的变异信息。第一排序轴与环境因子变量NO3-N(-0.3078)负相关,与P04-P(O.741 1)、NH4+-N(0.684 3)、TN(0.6266)、TP(0.546 6)、Chl a(0.385 4)和NO2-—N(0.085 2)和正相关,第二排序轴与环境因子变量Chl a(-0.7624)、NO2—N(-O.697 8)、NH4+一N(-0.633 4)、POt—P(-0.512 9)和TN(-0.464 9)负相关,与TP(0.653 3)和NO3一N(0.229 6)和正相关。说明影响7月份水体细菌群落碳源代谢功能环境因子主要是:Chl a、PO4-P、NO2-N、NH4+-N和TP,其中Chl a主要对AWCD(正相关)、phen(正相关)和carh(负相关)等产生影响,PO4-P和NH4+一N主要对AWCD(正相关)和poly(正相关)等产生影响,N02一N主要对Shannon(正相关)和phen(正相关)等产生影响,TP主要对S-E(正相关)、McIntosh(正相关)、Simpson(正相关)、carx(正相关)和phen(负相关)等产生影响。
8月,前两个排序轴的特征值为55%和25.1%,共能解释80.1%的变异信息,第一排序轴与环境因子变量NH4+-N(-0.1984)负相关,与NO3-N(O.935 5)、NO2
一N(0.925 0)、TP(0.7290)、TN(0.5475)、P04一P(0.5353)和Chla(0.0236)正相关。第二排序轴与环境因子变量Chl a(-0.011 6)负相关,与NH4+-N(0.3986)、PO4-P(0.2761)、TN(0.0876)、NO3-N(0.0760)、TP(0.0527)和NO2-N(0.0278)正相关。说明影响8月份水体水体细菌群落碳源代谢功能的环境因子主要是:NO3-N、NO2-N、TP、TN和PO4-P。其中这些环境因子主要是对:Shannon(正相关)、S-E(正相关)、phen(正相关)和carx(负相关)等产生影响。
9月,前两个排序轴的特征值为63.6%和25.1%,共能解释88.7%的变异信息,第一排序轴与环境因子变量NO3-N(-0.0719)负相关,与PO4-P(0.3897)、NH4+-N(0.2790)、TP(0.2430)、NO2--N(0.1516)、TN(0.058 4)和Chl a(0.024 9)正相关。第二排序轴与环境因子变量TP(一0.688 3)、Chl a(-0.5295)、NO2--N(-0.3724)、TN(-0.3344)、NO3--N(-0.2877)、PO4--P(-0.1732)和NH4+-N(-0.0824)负相关。说明影响9月份水体水体细菌群落碳代谢功能的环境因子主要是:TP、Chl a、PO4-P和NO2-N。其中TP和Chl a主要对phen(正相关)、Simpson(正相关)、Shannon(正相关)、S-E(正相关)amin(负相关)和carx(负相关)等产生影响,PO4-P和NO2-N主要对carx(负相关)、poly(负相关)、amin(负相关)和AWCD(负相关)等产生影响。
3.讨论
3.1碳氦比调控下虾、蟹混养系统水体中细菌群落的Biolog分析
Biolog方法对细菌群落多样性的检测是基于养殖环境内细菌对不同碳源利用程度的差异来实现的,通过检测ECO板吸光度的变化来评价不同环境区系细菌群落的生理特性和结构特征,还可以在空间及时间尺度上研究细菌群落的变化,具有易操作、灵敏度高、分辨能力强、数据丰富其可以直观的反应细菌群落结构的总体活性的优点。本实验利用该方法研究发现虾蟹混养系统不同处理水体中细菌群落对碳源总量和不同类型碳源的利用随养殖实验的延长差异越来越显著,这说明细菌群落对不同类型碳源利用喜好和对总碳源的代谢能力存在差异,反映了碳源和菌制剂的添加促使养殖水体中细菌群落的结构组成产生差异。这可能是:在养殖水体中添加碳源,调节碳氮比可以促进异养细菌的生长引,添加芽孢杆菌可以提高水体对碳源代谢活性和利用能力,添加硝化细菌或EM菌可以净化水质,但硝化细菌在添加一段时间后,会抑制了环境水体中异养细菌的生长繁殖,影响对碳源的代谢活性,这与本实验研究结果相似,说明养殖系统中添加碳源和菌制剂,影响了养殖环境中细菌群落的多样性。
Biolog的主成分分析可以用来直观地表述细菌群落代谢特征,是表征细菌群落功能多样性的重要手段,其分布的空间位置用来反映细菌群落对碳源底物的利用能力。本实验研究发现,虾蟹混养系统不同处理水体中细菌群落对碳源的利用能力差异显著(P<0.05)。7月,在分析图中分布均匀,说明不同处理间差异不显著,随着时间的推移,差异越来越显著。9月,不同处理间在分析图间的位置差异明显,其中处理组C3和C4在分析图中的位置与其他处理组显著不同,处理C3对六大碳源利用强度在总体上显著高于其他处理组(P<0.05),而处理组C4则对六大碳源的吸收水平较低稍高于对照组CO。这说明碳源和菌制剂的添加促使水体细菌群落不断发生动态变化直到达到稳定状态,最终导致C3组细菌群落组对碳源的代谢能力和强度最大,显著高于其它处理组。
细菌群落多样性指数不仅在总体上可以反应群落功能的动态变化,还是反映群落丰富度、均匀度和优势度的重要手段,但采用不同的多样性指数,会出现不同的研究结果l_3。。。本实验研究发现,不同处理水体细菌群落的McIntosh指数和Simpson指数随着养殖实验的进行呈现上升趋势,但八、九月相差不大,而Shannon指数和S-E均匀度指数均呈现先下降后上升的变化趋势。7月,处理组C3的Simpson指数和Shannon指均较低,而McIntosh指数和S-E均匀度指数则处于中间水平,处理组C4的McIntosh指数最低。8月,McIntosh指数在处理C3组中显著高于其它处理组,处理组C4组位于中间水平,其它多样性指数相差不大。9月,处理组C3中各类多样性指数均为最高,而处理组C4的McIntosh指数最低,Simpson指数和Shannon与对照组CO相差不大均显著低。这表明处理组C3细菌群落的空间多样性、物种丰富度和均匀度均较高,而C4组正好相反,这可能是处理组C4中存在明显的优势菌群,影响了其细菌群落的多样性和均匀度,从而影响了对底物碳总量和不同类型的利用。
3.2碳氦比调控下虾蟹混养系统水体中细菌群落碳代谢功能与环境因子的关系
本实验采用Biolog微平板培养法与冗余分析(RDA)进行相关分析,研究表明菌碳比调控下虾蟹混养系统水体中细菌群落碳代谢功能与养殖环境中理化因子间具有很好的相关性。结果显示,影响细菌群落碳代谢功能的主要环境因子7月份为:Chl a、PO4-P、NO2—N、NH4+-N,8月份为:NO3--N、NO2--N、TP、TN和PO4--P,9月份为:TP、Chl a、PO4-P和NO2--N。可以看出,N元素为养殖期间的主要影响因子,Chl a为养殖初期的主要影响因子,P元素为养殖中、后期的主要影响因子。这可能是在养殖实验的初期,水体中碳源含量相对较低,细菌的生长繁殖需要的碳源主要依赖于浮游植物释放的有机物;随着碳源的添加,养殖环境中含有充足的有机碳源,这时期环境内的NH4+-N(提供了55%~99%总氮吸收)等无机氮盐,可以直接影响了细菌的生长和代谢,也可以通过影响浮游细菌的捕食者来间接影响细菌种类的多样性;养殖水体中P元素(TP和PO4--P)具有溶解性和流动性较低的特点,而碳源的添加,使得TP和PO4--P相对含量进一步降低,因此P元素成为养殖中、后期的主要限制因子。
