电化学活性微生物在微生物燃料电池阳极中的应用
张逸驰 蒋昭泓 刘颖
摘 要 微生物燃料电池是一种利用电化学活性微生物催化降解有机物的同时产生电能的新型产能装置。阳极电化学活性微生物特征及催化活性是影响微生物燃料电池产电效率的关键因素之一。本文对应用于阳极上的电化学活性微生物的富集、来源、菌种的生理生化特征及产电能力等进行了综述。
关键词 微生物燃料电池; 电化学活性微生物; 异化铁还原菌; 阳极呼吸菌; 评述
1 引 言
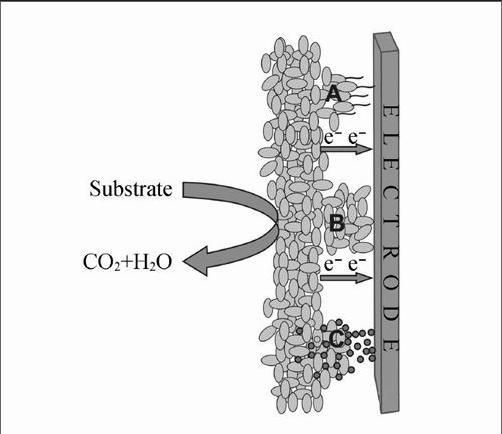
微生物燃料电池(Microbial fuel cell, MFC)可利用污水中的有机物、人类和动物排出的废物、农作物的废弃秸杆等发电,具有降解污染物的同时进行产能的双重功能,作为一种新能源技术已引起中外研究者们的极大兴趣[1~5],迄今已有百余年历史。由于输出功率较低,尚未实现有意义的大规模实际应用。近年来, MFC的输出功率已提高3~4个数量级[4,6,7]。在MFC的研究中,对用于阳极的产电微生物研究较为集中,富集在阳极上的电化学活性微生物将有机物氧化用于代谢过程中,产生的电子经过外电路传递给阴极,在阴极与电子受体结合,这些氧化还原反应使产生的电子不断传递并在外电路中形成电流[8,9]。在目前的报道中,MFC阳极中用于产生电流的微生物有几种名词,如电化学活性微生物、胞外产电菌及阳极呼吸菌(Electrochemically active bacteria, exoelectrogenic bacteria and anode respiring bacteria)等。电极表面性质和结构影响细菌的产电机理和产电能力,本文中统称这类产电的微生物为电化学活性微生物。由于各物质在电极上发生反应时表现出各自不同的电化学活性特征,以及不同产电微生物与电极间表现出电子传递特征的差异性[10],目前发现的微生物与电极间电子传递主要有纳米导线[2]、细胞外膜蛋白[11]及利用微生物自生中介体或外加中介体[12] 3种方式,完成微生物与电极间的电子转移。但常常几种方式同时作用,如图1所示。
这类电化学活性微生物在利用电子给体进行氧化的过程中,不同的微生物对于电子给体用于自身细胞物质的合成与用于产电的比率是不相同的;同一种微生物对不同电子给体的作用也是不同的。如有些微生物能将绝大多数电子给体氧化而用于产电,最高比率可达90%以上,有些微生物则将大部分有机物用于同化过程而只氧化少部分电子给体 (40%以下),可见电化学活性微生物在电极表面的催化活性是直接与菌种本身对底物利用的特性相关。因此,了解各产电微生物的生理生化特性、筛选优势的产电微生物及分析不同菌种间的协同作用,对于进一步深入揭示微生物在阳极表面的电子传递机制及提高电化学催化能力为加速微生物燃料电池的实际应用具有重要意义。
2 电化学活性微生物的来源、富集、分离及鉴定
有关电化学活性微生物在MFC中的研究主要有混合菌及纯菌种两类,纯菌种不但底物来源较窄而且操作要求非常严格,混合菌除了具有从周围环境中容易获得及对环境条件容易适应外,同时在操作上具有相对简单的特点,尤其是对底物的选择更加广泛,所以电化学活性混合菌在实际应用中表现出更加独特的优势。从环境中进行电化学活性微生物的富集培养可采用三电极体系的半电池装置或应用MFC装置体系,或者直接先筛选铁还原菌,再从中进一步筛选具有铁还原性的电化学活性微生物。另一方面,为更清晰地研究电化学活性微生物的具体产电机理,需要对群落中的优势菌株进行分离纯化,分离纯化后的菌株可作为理论模型菌利用其降解代谢的具体途径去探索其内部的产电机理。
2.1 电化学活性微生物来源、富集及群落特征
环境中存在着大量电化学活性微生物,广泛分布在工业废水包括乳业的粪肥、制作啤酒的废水、海底沉积物、厌氧活性污泥等自然环境中。目前,已有研究以厌氧颗粒污泥、消化污泥、生活污水以及牛胃液等作为接种源,成功启动微生物燃料电池,得到的电化学活性微生物群落表现出不同的群落特征及不同的产电能力。对于从不同来源和不同富集条件富集而来的微生物在种类、群落及产电能力上存在着很大差异。乙酸盐作为厌氧环境中含量最多最简单的有机酸,是MFC中最常用的底物,在以乙酸钠为底物富集到的MFC微生物群落中变形菌门(Proteobacteria)占多数,其中大部分为地杆菌属,地杆菌是阳极电化学活性微生物群落中的主要微生物[13]。另外,以不同有机酸为底物时,MFC的库伦效率明显不同,如醋酸盐、丙酸盐和丁酸盐为底物时,库仑效率分别为72.3%,36.0% 和43.0%[14]。葡萄糖也是MFC中常用底物之一,其中从美国Oyster Bay的厌氧沉积物中富集分离到的R. ferrireducens能直接将葡萄糖氧化为二氧化碳,但对大多数微生物而言,在以葡萄糖作为底物的MFC中,不同微生物降解葡萄糖时会产生不同种类的副产物,副产物对电池的功率密度和库仑效率也有影响,从而导致阳极上的电化学活性微生物种类也具有多样性。复杂有机物纤维素在自然界中普遍存在,已有利用瘤胃微生物作阳极生物催化剂与周期性添加纤维素作为底物成功使MFC运行2个多月的先例, 其中Clostridium spp.是附着生长的优势菌,当加入不同的底物时,所富集到的菌种也是不同的[15]。而在Rezaei等[16]设计的U形管MFC,同样以瘤胃微生物和纤维素分别为催化剂和为底物,分离到的可以利用纤维素产电的微生物是Enterobacter cloacae。 同样,从海底沉积物中及从河水中直接富集到的菌群中也有很大差别[13,17]。可见,所富集的菌群组成特征依赖于菌群的来源及富集环境。
对菌落的分析可采用传统的Sanger测序技术,但针对该技术检测通量低和检测灵敏度不高等问题,已有研究者采用其它联用技术提高测试性能。例如DGGE方法,具有可靠、可重复、快速等特点,已成为分析微生物群落的多样性及动态变化的重要工具,为目前分析MFC中微生物群落结构的常用方法[18]。基因文库分析法也是常用的分子生物学分析技术,进行16S rDNA克隆文库构建,获取更为精细的微生物群落结构,进而获得特定环境中的细菌群落结构和细菌多样性信息。通过构建16S rDNA克隆文库,已经取得以厌氧污泥为接种源,,并以麦秸为底物而富集的电化学活性微生物群落结构[18]。近来对菌落的群落分析出现了更为先进的高通量测序技术,该方法可以更加准确地对微生物群落进行分析,已逐渐成为目前研究微生物种群最先进的分子生物学技术。该方法已多次用于分析MFC中所利用的电化学活性微生物群落结构特征[19]。
2.2 电化学活性微生物菌种的分离与鉴定
对于表现出电化学活性的微生物群落可以进一步进行单一菌种的分离和鉴定工作,目前分离电化学活性微生物主要的应用方法有: 稀释涂布平板法[20]、生物激光印刷法(BioLP) [21]、U形管式的减绝-稀释培养法(Dilution-to-extinction)[22]等。
分离纯菌种的经典的稀释涂布平板方法可用于从三电极电化学体系或MFC装置富集到的群落中进行具有电化学活性的纯菌种的分离。该方法简单,易于操作,但由于这种方法很难直接用于从总菌落的稀释液中寻找产电菌,所以对于分离电化学活性菌有一定的困难。但是若欲分离具有铁还原能力的产电菌,可采取在培养基中加入柠檬酸铁,筛选需要Fe(Ⅲ)进行生长的电化学活性微生物,淘汰不具铁还原能力的菌种,进而提高选择到具有铁还原能力的电化学活性菌的机率。平板划线分离法与稀释平板法相比,操作较简单,但直接应用也存在对产电菌没有选择性的问题。减绝-稀释法的U型管法是最近发现的一种新的分离电化学活性菌的有效方法[23],具体过程是采用一个直型和一个弯曲型玻璃厌氧管分别作阳极和阴极室,将从电极收集的混合菌连续稀释后,重新接入MFC中,混合菌体在稀释到一定倍数时,群落中的劣势电化学活性菌种会被淘汰,从而使其中优势菌种在电极上得到单一培养,这样使分离到单一优势菌成为可能。这种方法避免了传统分离方法的冗长过程,对得到具有优势的电化学活性菌简单有效。该方法是根据菌株的电化学活性能力,而不是还原铁的能力将不同菌株分离,所以可以克服采用铁还原筛选法中丢失无还原铁能力的电化学活性菌的问题[24]。
对于直接分离得到的菌株,可采用16S rDNA方法进行微生物物种鉴定,通过对细菌基因组中的16S rDNA进行克隆测序,最后在Genbank数据库中进行核酸序列比对及大量的生理生化实验进行细菌鉴定。近年来,科研者分离出的具有电化学活性和产电功能的微生物种类很广,包括原核细菌、绿藻及真核酵母菌[25]。但是,总的来说目前已发现的电化学活性菌的种类还很有限,对这些微生物与电极间电子转移机理的认识还需要做更深入的工作。
3 电化学活性微生物的特征及产电能力
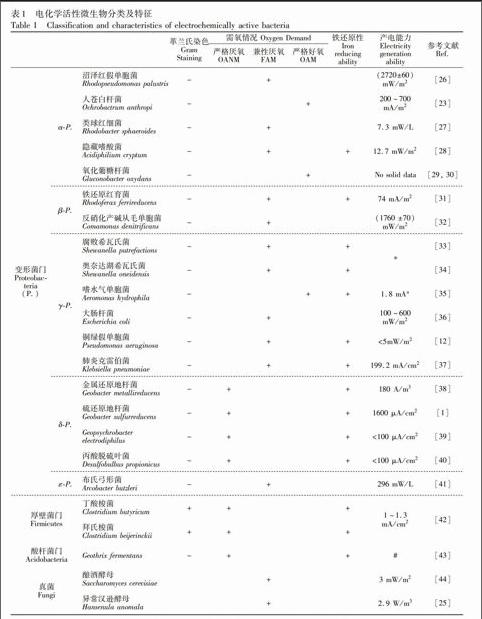
目前,从微生物燃料电池中分离出的电化学活性微生物主要以细菌为主,分别为变形菌门、厚壁菌门和酸杆菌门(见表1)。变形菌门是目前研究中发现电化学活性微生物最多的一门,且发现绝大多数产电菌为异化金属还原菌,并且多数为革兰氏阴性菌,其中兼性厌氧菌和严格厌氧菌偏多。Shewanella和Geobacter是最早研究和研究得较多的两个属,常用来作为研究微生物与电极间电子机理的模式菌。此外,真菌门的酵母菌也发现具有产电能力。第一个发现的不需添加介体的产电的微生物是异化Fe还原菌Shewanella putrefaciens[45],属γ-变形菌纲,但S. putrefaciens不能将底物彻底氧化,而能与电极间直接进行电子转移的δ-变形菌纲中的G. sulfurreducens可将醋酸钠彻底氧化为二氧化碳[13]。对Fe的还原能力及对氧的耐受性常常是菌种的主要特征,已发现的电化学活性微生物按其对Fe的还原能力可分为严格厌氧、兼性厌氧及专性好氧的异化铁还原菌或非异化铁还原菌5类,下面分别简述这几类主要电化学活性菌的生理生化及产电特征。
3.1 严格厌氧铁还原菌
铁元素是地壳中含量第四高的元素,与生物地球化学循环及土壤中许多重要元素转化密切相关,它对促进土壤物质循环具有重大的环境意义,其中铁还原微生物在元素的生物地球化学循环中扮演着重要角色。已有报道以固体铁氧化物为电子受体时细菌的电子转移过程与以固体电极为电子受体时有相似的机理,尽管目前对这些机理的认识还不是很清楚,但这已为进一步揭示电化学活性微生物与电极间的电子转移机制有很大帮助。目前发现的严格厌氧菌中能还原铁,且同时能以固体电极为电子受体的菌,主要分布在变形菌门中的δ-变形菌纲、厚壁菌门中的梭菌纲和酸杆菌门中的全噬菌纲中。其中,硫还原地杆菌(Geobacter sulfurreducens)和金属还原地杆菌(Geobacter metallireducens)是目前研究得较多的电化学活性菌种。其所形成的电化学活性微生物膜在底物不存在时通过循环伏安测试可以观察到两对明显的氧化还原峰(相对于Ag/AgCl电极,式电位分别为 Symbolm@@ 376和 Symbolm@@ 295 mV),在底物存在时表现出典型“S”形的对底物的催化特征[1,46]。
3.1.1 变形菌门中的δ-变形菌纲 (1)硫还原地杆菌(Geobacter sulfurreducens)属于Geobacteraceae科,地杆菌属(Geobacter),为革兰氏阴性菌,专性厌氧,电子供体较少,仅能以乙酸和氢气为电子供体,Fe、S、Co-EDTA、延胡索酸和苹果酸为电子受体,多分布于污泥中。G. sulfurreducens是最早报道的厌氧条件下以电极为电子受体完全氧化电子供体的微生物。由于G. sulfurreducens可在电极表面附着生长,细胞间又可形成多层细胞组成的厚度达50 μm厚的生物膜,它是研究具有电化学活性微生物膜的模式菌种,通过循环伏安法已证实该微生物膜具有较高的电化学活性(1600 μA/cm2)[1]。G. sulfurreducens的
全基因组的序列信息已阐明,可作为模式菌研究细胞与电极间电子传递机制。(2)金属还原地杆菌(Geobacter metallireducens)属地杆菌科(Geobacteraceae),革兰氏阴性菌, 专性厌氧,能还原铁、锰及铀等来降低放射性元素的污染,以氧化铁为电子受体时能氧化有机酸为二氧化碳及氧化短链脂肪酸、乙醇和苯等,具有降低或消除有害污染物毒性的能力,该菌可与电极间直接进行电子交换,产电能力较好(180 A/m3)[38]。(3)Geopsychrobacter electrodiphilus属于Geobacteraceae科,为革兰氏阴性菌,在MFC 中能彻底氧化乙酸、苹果酸、延胡索酸和柠檬酸等产电,其具有在低温海底环境中生长的优势使它更适合用于发展海水沉积型MFC[39]。(4)丙酸脱硫叶菌(Desulfoblbus propionicus)属于Desulfobulbaceae科,脱硫球茎菌属(Desulfoblbus),革兰氏阴性杆状菌,但通常用于MFC中产电效率较低(130 mA/L) [47]。
3.1.2 厚壁菌门中的梭菌纲(有电化学活性的革兰氏阳性菌) 丁酸梭菌(Clostridium butyricum)属于梭菌属,菌体梭状,革兰氏阳性,严格厌氧。C. butyricum EG3是首次报道的能利用淀粉等复杂多糖产电的革兰氏阳性菌株,革兰氏阳性菌与电极间的电子传递机理是否与革兰氏阴性菌相同还有待于深入研究[48]。此外,同属的C. beijerinckii(拜氏梭菌)也可以利用淀粉、糖蜜、葡萄糖和乳酸等产电。
3.1.3 酸杆菌门中的全噬菌纲(Holophagae) Geothrix fermentans属于全噬菌科(Holophagaceae),Geothrix属,严格厌氧,革兰氏阴性菌。G. fermentans是从石油污染的铁还原环境中分离出的严格厌氧菌,它以电极为唯一电子受体时,能够彻底氧化乙酸、琥珀酸、苹果酸、乳酸、丙酸等简单有机酸,但产电能力较低[43]。
3.2 兼性厌氧铁还原菌
3.2.1 变形菌门中的α-变形菌纲 隐藏嗜酸菌(Acidiphilium cryptum)属于Rhodospirillales目、Acetobacteraceae科、Acidiphilium属。Acidiphilium cryptum是从矿物污水的沉积物中分离出的一株革兰氏阴性细菌,A.cryptum是首次报道的可以在酸性条件下催化MFC的电化学活性微生物[28],但并不能直接将电子传递到电极上,所以在没有中间介体时无法产电。
3.2.2 变形菌门中的β-变形菌纲 铁还原红育菌(Rhodoferax ferrireducens)属于丛毛单胞菌科、红育菌属,革兰氏阴性,兼性厌氧。最适生长温度为25~30 ℃。在以Fe3+为电子受体时,可以将葡萄糖彻底转化为CO2获得能量生长,该菌是最早报道的以电极为电子受体时能直接彻底氧化葡萄糖的电化学活性微生物[31],是发展微生物燃料电池很有潜力的菌种。
3.2.3 变形菌门中的γ-变形菌纲 (1)希瓦氏菌(Shewanella)属于希瓦氏菌科(Shewanellaceae),革兰氏阴性、兼性厌氧。希瓦氏菌是研究MFC的模式菌属,该属可以将产生的电子直接传递到阳极。S. putrefactions IR-1是最早分离出来的能直接将电子传递给电极的电化学活性微生物,能还原铁[49,50]。而S. oneidensis DSP10是最早发现的可在有氧条件下产电的菌种,在好氧条件下能将乳酸氧化成CO2并产电,另一菌株S. oneidensis MR-1可以还原Fe和Mn(Ⅳ)的氧化物,是研究电化学活性微生物与电极间电子传递机理时常用的模式电化学活性微生物,已经发现该菌株可以分泌核黄素(Riboflavin),是该菌与电极间电子交换的不可缺少的一种媒介体[[49]。(2)铜绿假单胞菌(Pseudomonas aeruginosa) 属于假单胞菌科 (Pseudomona-daceae),革兰氏阴性杆菌,兼性好氧,P. aeruginosa在产电的同时能够产生绿脓菌素(Pyocyanin),并作为自身和其他菌种与电极间的电子传递介体而提高产电能力,该菌是最早被报道的自身能产生电子介体的微生物[51]。但由于绿脓菌素与其它人为添加的电子介体一样具有毒性,所以它并不是用于MFC中的理想电化学活性微生物。(3)肺炎克雷伯菌(Klebisella pneumonia) 属于肠杆菌科 Enterobacteriaceae,革兰氏阴性菌,在厌氧环境下能以电极为电子受体时在阳极上形成的生物膜可直接催化氧化多种有机物而产电,产电性能较好[37]。
3.3 好氧异化铁还原菌的变形菌门中的γ-变形菌纲
Aeromona shydrophilia(嗜水气单胞菌)属于Aeromonadaceae科,Aeromonas属,革兰氏阴性菌,由于其致病性,不宜用于MFC研究[52]。
3.4 兼性厌氧非铁还原菌
3.4.1 变形菌门中的α-变形菌纲 (1)类球红细菌(Rhodobacter sphaeroides),属于Rhodobacterales目、Rhodobacteraceae科、Rhodobacter属,光合细菌,为革兰氏阴性菌。在好氧黑暗条件下可以利用乙酸、柠檬酸、酒石酸、甘露醇、甲醇和乙醇等;在黑暗厌氧条件下,可以利用丙酮和糖类,但发酵能力微弱,此菌多分布于光照的死水中[53]。该菌用于光合生物燃料电池时在负载高时输出能量较低[27]。(2)沼泽红假单胞菌(Rhodopseudomonas palustris)属于慢生根瘤菌科(Bradyrhizobiaceae),革兰氏阴性。该菌最佳的生长方式为光能异养, R. palustris是最早报道具有电化学活性的α-变形菌,它能利用甲酸盐、丁酸盐、丙酸盐、戊酸盐、乳酸盐、延胡索酸盐、乙醇、甘油、酵母膏以及无机物硫代硫酸盐等多种底物产电,其中利用乙酸盐产电的功率输出密度最高[54]。最近,Logan研究组发现R. palustris DX-1菌株除了具有广泛的产电底物外,还具有更高的产电能力,用该菌催化的MFC最大产电功率输出远高于利用混合菌群[26]。
3.4.2 变形菌门中的β-变形菌纲 Comamonas denitrificans是一种反硝化细菌[55],属于Burkholderiales科,Comamonas属,革兰氏阴性,兼性厌氧。该菌可以利用延胡索酸、丙二酸、丙酮酸、乳酸以及乙酸为电子给体,是目前发现的唯一能产电的反硝化细菌,由于在硝酸盐存在时,在产电的同时可以产生氮气,所以可以维持阳极区的厌氧环境。
3.4.3 变形菌门中的γ-变形菌纲 大肠杆菌(Escherichia coli)属于肠杆菌科(Enterobacteriaceae),埃希菌属(Escherichia),革兰氏阴性菌,兼性厌氧,具有呼吸和发酵两种代谢类型。一般使用大肠杆菌时需要加入适合的介体如中性红才能在MFC中产电,有报道其最大输出功率为100 mW/m2 [36]。另外,也有报道E. coli K12可利用葡萄糖直接产电,产生的最大功率为600 mW/m2[36]。
3.5 好氧非铁还原菌的变形菌门中的α-变形菌纲(α-Proteobacteria)
(1)人苍白杆菌(Ochrobactrum anthropi)属于布鲁菌科(Brucellaceae),苍白杆菌属(Ochrobactrum),革兰氏阴性,专性好氧。 O. anthropi能利用广泛的底物,具有多种代谢途径,需要注意的是该菌是条件致病菌。O. anthropi YZ-1是Logan研究组首次利用减绝-稀释的U型管法分离出来的菌株[23]。(2)氧化葡糖杆菌(Gluconobacter oxydans)属于醋酸杆菌科(Acetobacteraceae),葡糖杆菌属(Gluconobacter),革兰氏阴性菌。该菌是由Lee在2002年首次报道,以电极为其电子受体在介体存在时利用MFC产电的特征来检测细菌对不同碳源的响应能力,但该菌产电能力较低[57]。
3.6 真菌界
近年来,在真菌界也发现一些具有电化学活性的微生物[58]。
3.6.1 酿酒酵母(Saccharomyces cerevisiae) 酿酒酵母菌属于真菌界 (Fungi)、酵母科(Saccharomycetaceae),兼性厌氧,可以葡萄糖作为底物及以亚甲基蓝作为中间介体进行产电。由于以酿酒酵母制备的MFC具有适用范围广和便于机理研究等方面的优势,该菌目前也成为MFC中的研究热点[59]。
3.6.2 异常汉逊酵母(Hansenul aanmala) 该酵母属于真菌界 (Fungi),毛霉科(Mucoraceae),兼性厌氧。 H.anmala是最早报道的可以产电的真菌,其产电不需要加入中间介体,可直接将电子传递到电极上[60]。该酵母可利用葡萄糖产电,但与其它产电细菌相比,酵母产电能力较低。
3.7 影响电化学活性微生物产电能力的主要因素
对于MFC的阳极,分布在细菌和真菌中的电化学活性微生物具有丰富的物种多样性,目前随着研究的不断深入仍在不断增加,这些已发现的不同菌株表现出明显不同的产电特性。尽管表1中所列出的不同菌株产生的电流或功率来源于一篇文献,实际上对于不同工作中的数据不能直接对比,但考虑到每个实验条件及操作方法,各数值间已表现出普遍性规律,如地杆菌产电能力较高,革兰氏阳性产电菌株较少,且产电能力也较弱。与传统燃料电池相比,目前所有菌株的产电能力仍极低。由于胞内的电子从胞内跨膜传递到胞外固体电极的过程中要将有机物在以电极为电子受体时进行氧化同时释放出电子和质子,释放出的电子要不断地由外膜传递到电极上进而进入外电路中。其中有些特定的菌株(如地杆菌属)具有内在独特的呼吸链表出对特定有机物的利用及向细胞外膜的电子传递能力较高,从而表现出较好的产电能力。可见,不断优化菌种并根据这些产电的优势菌株生产工程菌是提高电化学活性微生物产电能力的内在因素。另外,电极材料的结构和表面性质也是影响微生物产电的重要因素,多孔及石墨烯材料[6]为电子受体时, 产电菌已表现出较高的电化学催化性能。
4 MFC的应用展望
MFC将生物可降解的物质直接转化为电能,虽然MFC在电能输出方面目前尚无竞争优势,但是在很多方面仍有很好的应用前景。如利用电化学活性微生物构建的MFC或Microbial electrolysis fuel cell (MEC)平台,已在污水处理、生物修复、制取生物燃料及用于生物传感器等诸多方面显示出极大的开发应用前景[61~64]。尤其是利用MFC的电量与底物BOD浓度之间的线性关系,作为生物传感器可实现污水在线监测,同时克服传统BOD5方法耗时长、过程繁琐及不能在线检测的不足[64],利用优化的电化学活性微生物可进一步提高对污水中BOD监测的灵敏度及缩短响应时间,构建低成本和高性能的MFC型BOD传感器是发展污水监测的有潜力的方向。另外,MFC在[5]及二氧化碳还原[65]等方面的应用研究也在不断开发。但目前MFC的成本较高、输出功率仍不理想,微生物与电极间的电子转移机理尚不清楚,所以还未真正实现大规模的实际应用。
References
1 Liu Y, Kim H, Franklin R, Bond D R. Energ. Environ. Sci., 2010, 3(11): 1782-1788
2 Reguera G, McCarthy K D, Mehta T, Nicoll J S, Tuominen M T, Lovley D R. Nature, 2005, ?435: 1098-1101
3 Pandit S, Khilari S, Roy S, Pradhan D, Das D. Bioresource Technology, 2014, ?166: 451-457
4 Park I H, Christy M, Kim P, Nahm K S. Biosensors & Bioelectronics, 2014, ?58: 75-80
5 Sabina K, Fayidh M A, Archana G, Sivarajan M, Babuskin S, Babu P A S, Krishnan K R, Sukumar M. Environ. Technol., 2014, ?35(17): 2194-2203
6 Zhang Y Z, Mo G Q, Li X W, Zhang W D, Zhang J Q, Ye J S, Huang X D, Yu C Z. Journal of Power Sources, 2011, 196(13): 5402-5407
7 Ren H, Torres C I, Parameswaran P, Rittmann B E, Chae J. Biosensor. Bioelectron., 2014, ?61: 587-592
8 Schrder U. Phys. Chem. Chem. Phys., 2007, ?9: 2619-2629
9 Dolch K, Danzer J, Kabbeck T, Bierer B, Erben J, Forster A H, Maisch J, Nick P, Kerzenmacher S, Gescher J. Bioresource Technology, 2014, ?157: 284-292
10 Lovley D R. Curr. Opin. Biotechnol., 2008, ?19(6): 564-571
11 Liu Y, Bond D R. Chemsuchem., 2012, ?5: 1047-1053
12 Rabaey K, Boon N, Hofte M, Verstraete W. Environ. Sci. Technol., 2005, ?39(9): 3401-3408
13 Bond D R, Holmes D E, Tender L M, Lovley D R. Science, 2002, ?295(5554): 483-485
14 Chae K J, Choi M J, Lee J W, Kim K Y, Kim I S. Bioresource Technology, 2009, ?100(14): 3518-3525
15 Rismani-Yazdi H, Christy A D, Dehority B A, Morrison M, Yu Z, Tuovinen O H. Biotechnology and Bioengineering, 2007, ?97(6): 1398-1407
16 Rezaei F, Xing D F, Wagner R, Regan J M, Richard T L, Logan B E. Appl. Environ. Microbiol., 2009, ?75(11): 3673-3678
17 Tender L M, Reimers C E, Stecher H A, Holmes D E, Bond D R, Lowy D A, Pilobello K, Fertig S J, Lovley D R. Nature Biotechnology, 2002, 20(8): 821-825
18 Zhang Y F, Min B K, Huang L P, Angelidaki I. Appl. Environ. Microbiol., 2009, ?75(11): 3389-3395
19 Toh H, Sharma V K, Oshima K, Kondo S, Hattori M, Ward F B, Free A, Taylor T D. Journal of Bacteriology, 2011, 193(22): 6411-6412
20 Wang A J, Sun D, Ren N Q, Liu C, Liu W Z, Logan B E, Wu W M. Bioresource Technology, 2010, ?101(14): 5733-5735
21 Ringeisen B R, Lizewski S E, Fitzgerald L A, Biffinger J C, Knight C L, Crookes-Goodson W J, Wu P K. Electroanalysis, 2010, ?22(7-8): 875-882
22 Zuo Y, Xing D F, Regan J M, Logan B E. Appl. Environ. Microbiol., 2008, ?74(10): 3130-3137
23 Pham C A, Jung S J, Phung N T, Lee J, Ghang L S, Kim B H, Yi H, Chun J. FEMS Microbiol. Lett., 2003, ?223: 129-134
24 Xing D, Cheng S, Logan B E, Regan J M. Appl. Environ. Microbiol., 2010, ?85(5): 1575-1587
25 Prasad D, Arun S, Murugesan A, Padmanaban S, Satyanarayanan R S, Berchmans S, Yegnaraman V. Biosensors & Bioelectronics, 2007, ?22(11): 2604-2610
26 Xing D, Zuo Y, Cheng S, Regan J M, Logan B E. Environ. Sci. Technol., 2008, ?42(11): 4146-4151
27 Rosenbaum M, Schroder U, Scholz F. Environ. Sci. Technol., 2005, ?39(16): 6328-6333
28 Borole A P, O′Neill H, Tsouris C, Cesar S. Biotechnology Letters, 2008, ?30(8): 1367-1372
29 Alferov S V, Tomashevskaya L G, Ponamoreva O N, Bogdanovskaya V A, Reshetilov A N. Russian Journal of Electro-chemistry, 2006, ?42(4): 403-404
30 Reshetilov A, Alferov S, Tomashevskaya L, Ponamoreva O. Electroanalysis, 2006, ?18(19-20): 2030-2034
31 Chaudhuri S K, Lovley D R. Nature Biotechnology, 2003, ?21(10): 1229-1232
32 Xing D F, Cheng S A, Logan B E, Regan J M. Appl. Environ. Microbiol., 2010, ?85(5): 1575-1587
33 Kim H J, Park H S, Hyun M S, Chang I S, Kim M, Kim B H. Enzyme and Microbial Technology, 2002, ?30(2): 145-152
34 Biffinger J C, Pietron J, Ray R, Little B, Ringeisen B R. Biosens. Bioelectron., 2007, ?22: 1672-1679
35 Pham C A, Jung S J, Phung N T, Lee J, Chang I S, Kim B H, Yi H, Chun J. FEMS Microbiology Letters, 2003, ?223(1): 129-134
36 Zhang T, Cui C, Chen S, Yang H, Shen P. Electrochem. Commun., 2008, ?10(2): 293-297
37 DENG Li-Fang, LI Fang-Bai, ZHOU Shun-Gui, HUANG De-Yin, NI Jin-Ren. Chinese Science Bulletin, 2009, ?(19): 2983-2987
邓丽芳, 李芳柏, 周顺桂, 黄德银, 倪晋仁. 科学通报, 2009, ?(19): 2983-2987
38 Call D F, Logan B E. Biosensors & Bioelectronics, 2011, ?26(11): 4526-4531
39 Holmes D E, Nicoll J S, Bond D R, Lovley D R. Appl. Environ. Microbiol., 2004, ?70(10): 6023-6030
40 Holmes D E, Bond D R, Lovley D R. Appl. Environ. Microbiol., 2004, ?70(2): 1234-1237
41 Fedorovich V, Knighton M C, Pagaling E, Ward F B, Free A, Goryanin I. Appl. Environ. Microbiol., 2009, ?75(23): 7326-7334
42 Niessen J, Schroder U, Scholz F. Electrochem. Commun., 2004, ?6(9): 955-958
43 Bond D R, Lovley D R. Appl. Environ. Microbiol., 2005, ?71(4): 2186-2189
44 Sayed E T, Tsujiguchi T, Nakagawa N. Bioelectrochem., 2012, ?86: 97-101
45 Kim B H, Kim H J, Hyun M S, Park D H. J. Microbiol. Biotechnol., ?1999, ?9(2): 127-131
46 Liu Y, Harnisch F, Fricke K, Sietmann R, Schroder U. Biosens. Bioelectron., 2008, ?24(4): 1006-1011
47 Niessen J, Schroder U, Harnisch F, Scholz F. Letters in Applied Microbiology, 2005, ?41(3): 286-290
48 Ren Z, Steinberg L M, Regan J M. Water Science and Technology, 2008, ?58(3): 617-622
49 Marsili E, Baron D B, Shikhare I D, Coursolle D, Gralnick J A, Bond D R. PNAS, 2008, ?105(10): 3968-3973
50 Kim H J, Park H S, Hyun M S, Chang I S, Kim M, Kim B H. Enzyme and Microbial Technology, 2002, ?30(2): 145-152
51 Rabaey K, Boon N, Siciliano S D, Verhaege M, Verstraete W. Appl. Environ. Microbiol., 2004, ?70(9): 5373-5382
52 Pham C A, Jung S J, Phung N T, Lee J, Chang I S, Kim B H, Yi H, Chun J. FEMS Microbiol. Lett., 2003, ?223(1): 129-134
53 Gomelsky M, Kaplan S. Journal of Bacteriology, ?1996, ?178(7): 1946-1954
54 ZHANG Ling-Hua, KUANG Zhe-Shi, CHEN Wei, ZHANG Bao-Ling. Journal of South China Normal University, ?2001, ?(4): 37-39
张玲华, 邝哲师, 陈 薇, ?张宝玲. 华南师范大学学报 (自然科学版) , 2001, ?4: 37-39
55 Gumaelius L, Magnusson G, Pettersson B, Dalhammar G. International Journal of Systematic and Evolutionary Microbiology, 2001, ?51(3): 999-1006
56 Lee S A, Choi Y, Jung S H, Kim S. Bioelectrochem., 2002, ?57(2): 173-178
57 Walker A L, Walker Jr C W. Journal of Power Sources, 2006, ?160(1): 123-129
58 YIN Yun, LIU Yi-Sheng, WANG Yi-Fei, LI Wei, ZHENG Xiao-Dong. Chinese Journal of Applied & Environmental Biology, 2010, ?16(3): 412-414
殷 赟, 刘宜胜, 王一非, 李 伟, 郑晓冬. 应用与环境生物学报, 2010, ?16, (3): 412-414
59 Prasad D, Arun S, Murugesan M, Padmanaban S, Satyanarayanan R, Berchmans S, Yegnaraman V. Biosensors and Bioelectronics, 2007, ?22(11): 2604-2610
60 Touch N, Hibino T, Nagatsu Y, Tachiuchi K. Bioresource Technology, 2014, ?158: 225-230
61 Bond D R, Holmes D E, Tender L M, Lovley D R. Science, 2002, ?295(5554): 483-485
62 Cheng S, Logan B E. PNAS, 2007, ?104(47): 18871-18873
63 Moon H, Chang I S, Jang J K, Kim K S, Lee J, Lovitt R W, Kim B H. J. Microbiol. Biotechnol., 2005, ?15(1): 192-196
64 Villano M, Aulenta F, Ciucci C, Ferri T, Giuliano A, Majone M. Bioresource Technology, 2010, 101(9): 3085-3090〖ZK)〗
Application of Electrochemically Active Bacteria as
Anodic Biocatalyst in Microbial Fuel Cells
ZHANG Yi-Chi, JIANG Zhao-Hong, LIU Ying*
(College of Life Sciences, Northwest A & F University, Yangling 711100, China)
Abstract Microbial fuel cell (MFC) is a novel device with the function to produce energy and degrade organic materials. The characteristics of anodic electrochemically active bacteria and catalytic activity are one of key factors to affect MFC performance. This review summarized the enrichment, source, taxonomy, physiological and biochemical characteristics and electricity production ability of electrochemically active bacteria.
Keywords Microbial fuel cell; Electrochemically active bacteria; Dissimilatory iron-reducing bacteria; Anode respiring bacteria; Review
(Received 19 July 2014; accepted 7 October 2014)
This work was supported by the National Natural Science Foundation of China (No.21375107)
摘 要 微生物燃料电池是一种利用电化学活性微生物催化降解有机物的同时产生电能的新型产能装置。阳极电化学活性微生物特征及催化活性是影响微生物燃料电池产电效率的关键因素之一。本文对应用于阳极上的电化学活性微生物的富集、来源、菌种的生理生化特征及产电能力等进行了综述。
关键词 微生物燃料电池; 电化学活性微生物; 异化铁还原菌; 阳极呼吸菌; 评述
1 引 言
微生物燃料电池(Microbial fuel cell, MFC)可利用污水中的有机物、人类和动物排出的废物、农作物的废弃秸杆等发电,具有降解污染物的同时进行产能的双重功能,作为一种新能源技术已引起中外研究者们的极大兴趣[1~5],迄今已有百余年历史。由于输出功率较低,尚未实现有意义的大规模实际应用。近年来, MFC的输出功率已提高3~4个数量级[4,6,7]。在MFC的研究中,对用于阳极的产电微生物研究较为集中,富集在阳极上的电化学活性微生物将有机物氧化用于代谢过程中,产生的电子经过外电路传递给阴极,在阴极与电子受体结合,这些氧化还原反应使产生的电子不断传递并在外电路中形成电流[8,9]。在目前的报道中,MFC阳极中用于产生电流的微生物有几种名词,如电化学活性微生物、胞外产电菌及阳极呼吸菌(Electrochemically active bacteria, exoelectrogenic bacteria and anode respiring bacteria)等。电极表面性质和结构影响细菌的产电机理和产电能力,本文中统称这类产电的微生物为电化学活性微生物。由于各物质在电极上发生反应时表现出各自不同的电化学活性特征,以及不同产电微生物与电极间表现出电子传递特征的差异性[10],目前发现的微生物与电极间电子传递主要有纳米导线[2]、细胞外膜蛋白[11]及利用微生物自生中介体或外加中介体[12] 3种方式,完成微生物与电极间的电子转移。但常常几种方式同时作用,如图1所示。
这类电化学活性微生物在利用电子给体进行氧化的过程中,不同的微生物对于电子给体用于自身细胞物质的合成与用于产电的比率是不相同的;同一种微生物对不同电子给体的作用也是不同的。如有些微生物能将绝大多数电子给体氧化而用于产电,最高比率可达90%以上,有些微生物则将大部分有机物用于同化过程而只氧化少部分电子给体 (40%以下),可见电化学活性微生物在电极表面的催化活性是直接与菌种本身对底物利用的特性相关。因此,了解各产电微生物的生理生化特性、筛选优势的产电微生物及分析不同菌种间的协同作用,对于进一步深入揭示微生物在阳极表面的电子传递机制及提高电化学催化能力为加速微生物燃料电池的实际应用具有重要意义。
2 电化学活性微生物的来源、富集、分离及鉴定
有关电化学活性微生物在MFC中的研究主要有混合菌及纯菌种两类,纯菌种不但底物来源较窄而且操作要求非常严格,混合菌除了具有从周围环境中容易获得及对环境条件容易适应外,同时在操作上具有相对简单的特点,尤其是对底物的选择更加广泛,所以电化学活性混合菌在实际应用中表现出更加独特的优势。从环境中进行电化学活性微生物的富集培养可采用三电极体系的半电池装置或应用MFC装置体系,或者直接先筛选铁还原菌,再从中进一步筛选具有铁还原性的电化学活性微生物。另一方面,为更清晰地研究电化学活性微生物的具体产电机理,需要对群落中的优势菌株进行分离纯化,分离纯化后的菌株可作为理论模型菌利用其降解代谢的具体途径去探索其内部的产电机理。
2.1 电化学活性微生物来源、富集及群落特征
环境中存在着大量电化学活性微生物,广泛分布在工业废水包括乳业的粪肥、制作啤酒的废水、海底沉积物、厌氧活性污泥等自然环境中。目前,已有研究以厌氧颗粒污泥、消化污泥、生活污水以及牛胃液等作为接种源,成功启动微生物燃料电池,得到的电化学活性微生物群落表现出不同的群落特征及不同的产电能力。对于从不同来源和不同富集条件富集而来的微生物在种类、群落及产电能力上存在着很大差异。乙酸盐作为厌氧环境中含量最多最简单的有机酸,是MFC中最常用的底物,在以乙酸钠为底物富集到的MFC微生物群落中变形菌门(Proteobacteria)占多数,其中大部分为地杆菌属,地杆菌是阳极电化学活性微生物群落中的主要微生物[13]。另外,以不同有机酸为底物时,MFC的库伦效率明显不同,如醋酸盐、丙酸盐和丁酸盐为底物时,库仑效率分别为72.3%,36.0% 和43.0%[14]。葡萄糖也是MFC中常用底物之一,其中从美国Oyster Bay的厌氧沉积物中富集分离到的R. ferrireducens能直接将葡萄糖氧化为二氧化碳,但对大多数微生物而言,在以葡萄糖作为底物的MFC中,不同微生物降解葡萄糖时会产生不同种类的副产物,副产物对电池的功率密度和库仑效率也有影响,从而导致阳极上的电化学活性微生物种类也具有多样性。复杂有机物纤维素在自然界中普遍存在,已有利用瘤胃微生物作阳极生物催化剂与周期性添加纤维素作为底物成功使MFC运行2个多月的先例, 其中Clostridium spp.是附着生长的优势菌,当加入不同的底物时,所富集到的菌种也是不同的[15]。而在Rezaei等[16]设计的U形管MFC,同样以瘤胃微生物和纤维素分别为催化剂和为底物,分离到的可以利用纤维素产电的微生物是Enterobacter cloacae。 同样,从海底沉积物中及从河水中直接富集到的菌群中也有很大差别[13,17]。可见,所富集的菌群组成特征依赖于菌群的来源及富集环境。
对菌落的分析可采用传统的Sanger测序技术,但针对该技术检测通量低和检测灵敏度不高等问题,已有研究者采用其它联用技术提高测试性能。例如DGGE方法,具有可靠、可重复、快速等特点,已成为分析微生物群落的多样性及动态变化的重要工具,为目前分析MFC中微生物群落结构的常用方法[18]。基因文库分析法也是常用的分子生物学分析技术,进行16S rDNA克隆文库构建,获取更为精细的微生物群落结构,进而获得特定环境中的细菌群落结构和细菌多样性信息。通过构建16S rDNA克隆文库,已经取得以厌氧污泥为接种源,,并以麦秸为底物而富集的电化学活性微生物群落结构[18]。近来对菌落的群落分析出现了更为先进的高通量测序技术,该方法可以更加准确地对微生物群落进行分析,已逐渐成为目前研究微生物种群最先进的分子生物学技术。该方法已多次用于分析MFC中所利用的电化学活性微生物群落结构特征[19]。
2.2 电化学活性微生物菌种的分离与鉴定
对于表现出电化学活性的微生物群落可以进一步进行单一菌种的分离和鉴定工作,目前分离电化学活性微生物主要的应用方法有: 稀释涂布平板法[20]、生物激光印刷法(BioLP) [21]、U形管式的减绝-稀释培养法(Dilution-to-extinction)[22]等。
分离纯菌种的经典的稀释涂布平板方法可用于从三电极电化学体系或MFC装置富集到的群落中进行具有电化学活性的纯菌种的分离。该方法简单,易于操作,但由于这种方法很难直接用于从总菌落的稀释液中寻找产电菌,所以对于分离电化学活性菌有一定的困难。但是若欲分离具有铁还原能力的产电菌,可采取在培养基中加入柠檬酸铁,筛选需要Fe(Ⅲ)进行生长的电化学活性微生物,淘汰不具铁还原能力的菌种,进而提高选择到具有铁还原能力的电化学活性菌的机率。平板划线分离法与稀释平板法相比,操作较简单,但直接应用也存在对产电菌没有选择性的问题。减绝-稀释法的U型管法是最近发现的一种新的分离电化学活性菌的有效方法[23],具体过程是采用一个直型和一个弯曲型玻璃厌氧管分别作阳极和阴极室,将从电极收集的混合菌连续稀释后,重新接入MFC中,混合菌体在稀释到一定倍数时,群落中的劣势电化学活性菌种会被淘汰,从而使其中优势菌种在电极上得到单一培养,这样使分离到单一优势菌成为可能。这种方法避免了传统分离方法的冗长过程,对得到具有优势的电化学活性菌简单有效。该方法是根据菌株的电化学活性能力,而不是还原铁的能力将不同菌株分离,所以可以克服采用铁还原筛选法中丢失无还原铁能力的电化学活性菌的问题[24]。
对于直接分离得到的菌株,可采用16S rDNA方法进行微生物物种鉴定,通过对细菌基因组中的16S rDNA进行克隆测序,最后在Genbank数据库中进行核酸序列比对及大量的生理生化实验进行细菌鉴定。近年来,科研者分离出的具有电化学活性和产电功能的微生物种类很广,包括原核细菌、绿藻及真核酵母菌[25]。但是,总的来说目前已发现的电化学活性菌的种类还很有限,对这些微生物与电极间电子转移机理的认识还需要做更深入的工作。
3 电化学活性微生物的特征及产电能力
目前,从微生物燃料电池中分离出的电化学活性微生物主要以细菌为主,分别为变形菌门、厚壁菌门和酸杆菌门(见表1)。变形菌门是目前研究中发现电化学活性微生物最多的一门,且发现绝大多数产电菌为异化金属还原菌,并且多数为革兰氏阴性菌,其中兼性厌氧菌和严格厌氧菌偏多。Shewanella和Geobacter是最早研究和研究得较多的两个属,常用来作为研究微生物与电极间电子机理的模式菌。此外,真菌门的酵母菌也发现具有产电能力。第一个发现的不需添加介体的产电的微生物是异化Fe还原菌Shewanella putrefaciens[45],属γ-变形菌纲,但S. putrefaciens不能将底物彻底氧化,而能与电极间直接进行电子转移的δ-变形菌纲中的G. sulfurreducens可将醋酸钠彻底氧化为二氧化碳[13]。对Fe的还原能力及对氧的耐受性常常是菌种的主要特征,已发现的电化学活性微生物按其对Fe的还原能力可分为严格厌氧、兼性厌氧及专性好氧的异化铁还原菌或非异化铁还原菌5类,下面分别简述这几类主要电化学活性菌的生理生化及产电特征。
3.1 严格厌氧铁还原菌
铁元素是地壳中含量第四高的元素,与生物地球化学循环及土壤中许多重要元素转化密切相关,它对促进土壤物质循环具有重大的环境意义,其中铁还原微生物在元素的生物地球化学循环中扮演着重要角色。已有报道以固体铁氧化物为电子受体时细菌的电子转移过程与以固体电极为电子受体时有相似的机理,尽管目前对这些机理的认识还不是很清楚,但这已为进一步揭示电化学活性微生物与电极间的电子转移机制有很大帮助。目前发现的严格厌氧菌中能还原铁,且同时能以固体电极为电子受体的菌,主要分布在变形菌门中的δ-变形菌纲、厚壁菌门中的梭菌纲和酸杆菌门中的全噬菌纲中。其中,硫还原地杆菌(Geobacter sulfurreducens)和金属还原地杆菌(Geobacter metallireducens)是目前研究得较多的电化学活性菌种。其所形成的电化学活性微生物膜在底物不存在时通过循环伏安测试可以观察到两对明显的氧化还原峰(相对于Ag/AgCl电极,式电位分别为 Symbolm@@ 376和 Symbolm@@ 295 mV),在底物存在时表现出典型“S”形的对底物的催化特征[1,46]。
3.1.1 变形菌门中的δ-变形菌纲 (1)硫还原地杆菌(Geobacter sulfurreducens)属于Geobacteraceae科,地杆菌属(Geobacter),为革兰氏阴性菌,专性厌氧,电子供体较少,仅能以乙酸和氢气为电子供体,Fe、S、Co-EDTA、延胡索酸和苹果酸为电子受体,多分布于污泥中。G. sulfurreducens是最早报道的厌氧条件下以电极为电子受体完全氧化电子供体的微生物。由于G. sulfurreducens可在电极表面附着生长,细胞间又可形成多层细胞组成的厚度达50 μm厚的生物膜,它是研究具有电化学活性微生物膜的模式菌种,通过循环伏安法已证实该微生物膜具有较高的电化学活性(1600 μA/cm2)[1]。G. sulfurreducens的
全基因组的序列信息已阐明,可作为模式菌研究细胞与电极间电子传递机制。(2)金属还原地杆菌(Geobacter metallireducens)属地杆菌科(Geobacteraceae),革兰氏阴性菌, 专性厌氧,能还原铁、锰及铀等来降低放射性元素的污染,以氧化铁为电子受体时能氧化有机酸为二氧化碳及氧化短链脂肪酸、乙醇和苯等,具有降低或消除有害污染物毒性的能力,该菌可与电极间直接进行电子交换,产电能力较好(180 A/m3)[38]。(3)Geopsychrobacter electrodiphilus属于Geobacteraceae科,为革兰氏阴性菌,在MFC 中能彻底氧化乙酸、苹果酸、延胡索酸和柠檬酸等产电,其具有在低温海底环境中生长的优势使它更适合用于发展海水沉积型MFC[39]。(4)丙酸脱硫叶菌(Desulfoblbus propionicus)属于Desulfobulbaceae科,脱硫球茎菌属(Desulfoblbus),革兰氏阴性杆状菌,但通常用于MFC中产电效率较低(130 mA/L) [47]。
3.1.2 厚壁菌门中的梭菌纲(有电化学活性的革兰氏阳性菌) 丁酸梭菌(Clostridium butyricum)属于梭菌属,菌体梭状,革兰氏阳性,严格厌氧。C. butyricum EG3是首次报道的能利用淀粉等复杂多糖产电的革兰氏阳性菌株,革兰氏阳性菌与电极间的电子传递机理是否与革兰氏阴性菌相同还有待于深入研究[48]。此外,同属的C. beijerinckii(拜氏梭菌)也可以利用淀粉、糖蜜、葡萄糖和乳酸等产电。
3.1.3 酸杆菌门中的全噬菌纲(Holophagae) Geothrix fermentans属于全噬菌科(Holophagaceae),Geothrix属,严格厌氧,革兰氏阴性菌。G. fermentans是从石油污染的铁还原环境中分离出的严格厌氧菌,它以电极为唯一电子受体时,能够彻底氧化乙酸、琥珀酸、苹果酸、乳酸、丙酸等简单有机酸,但产电能力较低[43]。
3.2 兼性厌氧铁还原菌
3.2.1 变形菌门中的α-变形菌纲 隐藏嗜酸菌(Acidiphilium cryptum)属于Rhodospirillales目、Acetobacteraceae科、Acidiphilium属。Acidiphilium cryptum是从矿物污水的沉积物中分离出的一株革兰氏阴性细菌,A.cryptum是首次报道的可以在酸性条件下催化MFC的电化学活性微生物[28],但并不能直接将电子传递到电极上,所以在没有中间介体时无法产电。
3.2.2 变形菌门中的β-变形菌纲 铁还原红育菌(Rhodoferax ferrireducens)属于丛毛单胞菌科、红育菌属,革兰氏阴性,兼性厌氧。最适生长温度为25~30 ℃。在以Fe3+为电子受体时,可以将葡萄糖彻底转化为CO2获得能量生长,该菌是最早报道的以电极为电子受体时能直接彻底氧化葡萄糖的电化学活性微生物[31],是发展微生物燃料电池很有潜力的菌种。
3.2.3 变形菌门中的γ-变形菌纲 (1)希瓦氏菌(Shewanella)属于希瓦氏菌科(Shewanellaceae),革兰氏阴性、兼性厌氧。希瓦氏菌是研究MFC的模式菌属,该属可以将产生的电子直接传递到阳极。S. putrefactions IR-1是最早分离出来的能直接将电子传递给电极的电化学活性微生物,能还原铁[49,50]。而S. oneidensis DSP10是最早发现的可在有氧条件下产电的菌种,在好氧条件下能将乳酸氧化成CO2并产电,另一菌株S. oneidensis MR-1可以还原Fe和Mn(Ⅳ)的氧化物,是研究电化学活性微生物与电极间电子传递机理时常用的模式电化学活性微生物,已经发现该菌株可以分泌核黄素(Riboflavin),是该菌与电极间电子交换的不可缺少的一种媒介体[[49]。(2)铜绿假单胞菌(Pseudomonas aeruginosa) 属于假单胞菌科 (Pseudomona-daceae),革兰氏阴性杆菌,兼性好氧,P. aeruginosa在产电的同时能够产生绿脓菌素(Pyocyanin),并作为自身和其他菌种与电极间的电子传递介体而提高产电能力,该菌是最早被报道的自身能产生电子介体的微生物[51]。但由于绿脓菌素与其它人为添加的电子介体一样具有毒性,所以它并不是用于MFC中的理想电化学活性微生物。(3)肺炎克雷伯菌(Klebisella pneumonia) 属于肠杆菌科 Enterobacteriaceae,革兰氏阴性菌,在厌氧环境下能以电极为电子受体时在阳极上形成的生物膜可直接催化氧化多种有机物而产电,产电性能较好[37]。
3.3 好氧异化铁还原菌的变形菌门中的γ-变形菌纲
Aeromona shydrophilia(嗜水气单胞菌)属于Aeromonadaceae科,Aeromonas属,革兰氏阴性菌,由于其致病性,不宜用于MFC研究[52]。
3.4 兼性厌氧非铁还原菌
3.4.1 变形菌门中的α-变形菌纲 (1)类球红细菌(Rhodobacter sphaeroides),属于Rhodobacterales目、Rhodobacteraceae科、Rhodobacter属,光合细菌,为革兰氏阴性菌。在好氧黑暗条件下可以利用乙酸、柠檬酸、酒石酸、甘露醇、甲醇和乙醇等;在黑暗厌氧条件下,可以利用丙酮和糖类,但发酵能力微弱,此菌多分布于光照的死水中[53]。该菌用于光合生物燃料电池时在负载高时输出能量较低[27]。(2)沼泽红假单胞菌(Rhodopseudomonas palustris)属于慢生根瘤菌科(Bradyrhizobiaceae),革兰氏阴性。该菌最佳的生长方式为光能异养, R. palustris是最早报道具有电化学活性的α-变形菌,它能利用甲酸盐、丁酸盐、丙酸盐、戊酸盐、乳酸盐、延胡索酸盐、乙醇、甘油、酵母膏以及无机物硫代硫酸盐等多种底物产电,其中利用乙酸盐产电的功率输出密度最高[54]。最近,Logan研究组发现R. palustris DX-1菌株除了具有广泛的产电底物外,还具有更高的产电能力,用该菌催化的MFC最大产电功率输出远高于利用混合菌群[26]。
3.4.2 变形菌门中的β-变形菌纲 Comamonas denitrificans是一种反硝化细菌[55],属于Burkholderiales科,Comamonas属,革兰氏阴性,兼性厌氧。该菌可以利用延胡索酸、丙二酸、丙酮酸、乳酸以及乙酸为电子给体,是目前发现的唯一能产电的反硝化细菌,由于在硝酸盐存在时,在产电的同时可以产生氮气,所以可以维持阳极区的厌氧环境。
3.4.3 变形菌门中的γ-变形菌纲 大肠杆菌(Escherichia coli)属于肠杆菌科(Enterobacteriaceae),埃希菌属(Escherichia),革兰氏阴性菌,兼性厌氧,具有呼吸和发酵两种代谢类型。一般使用大肠杆菌时需要加入适合的介体如中性红才能在MFC中产电,有报道其最大输出功率为100 mW/m2 [36]。另外,也有报道E. coli K12可利用葡萄糖直接产电,产生的最大功率为600 mW/m2[36]。
3.5 好氧非铁还原菌的变形菌门中的α-变形菌纲(α-Proteobacteria)
(1)人苍白杆菌(Ochrobactrum anthropi)属于布鲁菌科(Brucellaceae),苍白杆菌属(Ochrobactrum),革兰氏阴性,专性好氧。 O. anthropi能利用广泛的底物,具有多种代谢途径,需要注意的是该菌是条件致病菌。O. anthropi YZ-1是Logan研究组首次利用减绝-稀释的U型管法分离出来的菌株[23]。(2)氧化葡糖杆菌(Gluconobacter oxydans)属于醋酸杆菌科(Acetobacteraceae),葡糖杆菌属(Gluconobacter),革兰氏阴性菌。该菌是由Lee在2002年首次报道,以电极为其电子受体在介体存在时利用MFC产电的特征来检测细菌对不同碳源的响应能力,但该菌产电能力较低[57]。
3.6 真菌界
近年来,在真菌界也发现一些具有电化学活性的微生物[58]。
3.6.1 酿酒酵母(Saccharomyces cerevisiae) 酿酒酵母菌属于真菌界 (Fungi)、酵母科(Saccharomycetaceae),兼性厌氧,可以葡萄糖作为底物及以亚甲基蓝作为中间介体进行产电。由于以酿酒酵母制备的MFC具有适用范围广和便于机理研究等方面的优势,该菌目前也成为MFC中的研究热点[59]。
3.6.2 异常汉逊酵母(Hansenul aanmala) 该酵母属于真菌界 (Fungi),毛霉科(Mucoraceae),兼性厌氧。 H.anmala是最早报道的可以产电的真菌,其产电不需要加入中间介体,可直接将电子传递到电极上[60]。该酵母可利用葡萄糖产电,但与其它产电细菌相比,酵母产电能力较低。
3.7 影响电化学活性微生物产电能力的主要因素
对于MFC的阳极,分布在细菌和真菌中的电化学活性微生物具有丰富的物种多样性,目前随着研究的不断深入仍在不断增加,这些已发现的不同菌株表现出明显不同的产电特性。尽管表1中所列出的不同菌株产生的电流或功率来源于一篇文献,实际上对于不同工作中的数据不能直接对比,但考虑到每个实验条件及操作方法,各数值间已表现出普遍性规律,如地杆菌产电能力较高,革兰氏阳性产电菌株较少,且产电能力也较弱。与传统燃料电池相比,目前所有菌株的产电能力仍极低。由于胞内的电子从胞内跨膜传递到胞外固体电极的过程中要将有机物在以电极为电子受体时进行氧化同时释放出电子和质子,释放出的电子要不断地由外膜传递到电极上进而进入外电路中。其中有些特定的菌株(如地杆菌属)具有内在独特的呼吸链表出对特定有机物的利用及向细胞外膜的电子传递能力较高,从而表现出较好的产电能力。可见,不断优化菌种并根据这些产电的优势菌株生产工程菌是提高电化学活性微生物产电能力的内在因素。另外,电极材料的结构和表面性质也是影响微生物产电的重要因素,多孔及石墨烯材料[6]为电子受体时, 产电菌已表现出较高的电化学催化性能。
4 MFC的应用展望
MFC将生物可降解的物质直接转化为电能,虽然MFC在电能输出方面目前尚无竞争优势,但是在很多方面仍有很好的应用前景。如利用电化学活性微生物构建的MFC或Microbial electrolysis fuel cell (MEC)平台,已在污水处理、生物修复、制取生物燃料及用于生物传感器等诸多方面显示出极大的开发应用前景[61~64]。尤其是利用MFC的电量与底物BOD浓度之间的线性关系,作为生物传感器可实现污水在线监测,同时克服传统BOD5方法耗时长、过程繁琐及不能在线检测的不足[64],利用优化的电化学活性微生物可进一步提高对污水中BOD监测的灵敏度及缩短响应时间,构建低成本和高性能的MFC型BOD传感器是发展污水监测的有潜力的方向。另外,MFC在[5]及二氧化碳还原[65]等方面的应用研究也在不断开发。但目前MFC的成本较高、输出功率仍不理想,微生物与电极间的电子转移机理尚不清楚,所以还未真正实现大规模的实际应用。
References
1 Liu Y, Kim H, Franklin R, Bond D R. Energ. Environ. Sci., 2010, 3(11): 1782-1788
2 Reguera G, McCarthy K D, Mehta T, Nicoll J S, Tuominen M T, Lovley D R. Nature, 2005, ?435: 1098-1101
3 Pandit S, Khilari S, Roy S, Pradhan D, Das D. Bioresource Technology, 2014, ?166: 451-457
4 Park I H, Christy M, Kim P, Nahm K S. Biosensors & Bioelectronics, 2014, ?58: 75-80
5 Sabina K, Fayidh M A, Archana G, Sivarajan M, Babuskin S, Babu P A S, Krishnan K R, Sukumar M. Environ. Technol., 2014, ?35(17): 2194-2203
6 Zhang Y Z, Mo G Q, Li X W, Zhang W D, Zhang J Q, Ye J S, Huang X D, Yu C Z. Journal of Power Sources, 2011, 196(13): 5402-5407
7 Ren H, Torres C I, Parameswaran P, Rittmann B E, Chae J. Biosensor. Bioelectron., 2014, ?61: 587-592
8 Schrder U. Phys. Chem. Chem. Phys., 2007, ?9: 2619-2629
9 Dolch K, Danzer J, Kabbeck T, Bierer B, Erben J, Forster A H, Maisch J, Nick P, Kerzenmacher S, Gescher J. Bioresource Technology, 2014, ?157: 284-292
10 Lovley D R. Curr. Opin. Biotechnol., 2008, ?19(6): 564-571
11 Liu Y, Bond D R. Chemsuchem., 2012, ?5: 1047-1053
12 Rabaey K, Boon N, Hofte M, Verstraete W. Environ. Sci. Technol., 2005, ?39(9): 3401-3408
13 Bond D R, Holmes D E, Tender L M, Lovley D R. Science, 2002, ?295(5554): 483-485
14 Chae K J, Choi M J, Lee J W, Kim K Y, Kim I S. Bioresource Technology, 2009, ?100(14): 3518-3525
15 Rismani-Yazdi H, Christy A D, Dehority B A, Morrison M, Yu Z, Tuovinen O H. Biotechnology and Bioengineering, 2007, ?97(6): 1398-1407
16 Rezaei F, Xing D F, Wagner R, Regan J M, Richard T L, Logan B E. Appl. Environ. Microbiol., 2009, ?75(11): 3673-3678
17 Tender L M, Reimers C E, Stecher H A, Holmes D E, Bond D R, Lowy D A, Pilobello K, Fertig S J, Lovley D R. Nature Biotechnology, 2002, 20(8): 821-825
18 Zhang Y F, Min B K, Huang L P, Angelidaki I. Appl. Environ. Microbiol., 2009, ?75(11): 3389-3395
19 Toh H, Sharma V K, Oshima K, Kondo S, Hattori M, Ward F B, Free A, Taylor T D. Journal of Bacteriology, 2011, 193(22): 6411-6412
20 Wang A J, Sun D, Ren N Q, Liu C, Liu W Z, Logan B E, Wu W M. Bioresource Technology, 2010, ?101(14): 5733-5735
21 Ringeisen B R, Lizewski S E, Fitzgerald L A, Biffinger J C, Knight C L, Crookes-Goodson W J, Wu P K. Electroanalysis, 2010, ?22(7-8): 875-882
22 Zuo Y, Xing D F, Regan J M, Logan B E. Appl. Environ. Microbiol., 2008, ?74(10): 3130-3137
23 Pham C A, Jung S J, Phung N T, Lee J, Ghang L S, Kim B H, Yi H, Chun J. FEMS Microbiol. Lett., 2003, ?223: 129-134
24 Xing D, Cheng S, Logan B E, Regan J M. Appl. Environ. Microbiol., 2010, ?85(5): 1575-1587
25 Prasad D, Arun S, Murugesan A, Padmanaban S, Satyanarayanan R S, Berchmans S, Yegnaraman V. Biosensors & Bioelectronics, 2007, ?22(11): 2604-2610
26 Xing D, Zuo Y, Cheng S, Regan J M, Logan B E. Environ. Sci. Technol., 2008, ?42(11): 4146-4151
27 Rosenbaum M, Schroder U, Scholz F. Environ. Sci. Technol., 2005, ?39(16): 6328-6333
28 Borole A P, O′Neill H, Tsouris C, Cesar S. Biotechnology Letters, 2008, ?30(8): 1367-1372
29 Alferov S V, Tomashevskaya L G, Ponamoreva O N, Bogdanovskaya V A, Reshetilov A N. Russian Journal of Electro-chemistry, 2006, ?42(4): 403-404
30 Reshetilov A, Alferov S, Tomashevskaya L, Ponamoreva O. Electroanalysis, 2006, ?18(19-20): 2030-2034
31 Chaudhuri S K, Lovley D R. Nature Biotechnology, 2003, ?21(10): 1229-1232
32 Xing D F, Cheng S A, Logan B E, Regan J M. Appl. Environ. Microbiol., 2010, ?85(5): 1575-1587
33 Kim H J, Park H S, Hyun M S, Chang I S, Kim M, Kim B H. Enzyme and Microbial Technology, 2002, ?30(2): 145-152
34 Biffinger J C, Pietron J, Ray R, Little B, Ringeisen B R. Biosens. Bioelectron., 2007, ?22: 1672-1679
35 Pham C A, Jung S J, Phung N T, Lee J, Chang I S, Kim B H, Yi H, Chun J. FEMS Microbiology Letters, 2003, ?223(1): 129-134
36 Zhang T, Cui C, Chen S, Yang H, Shen P. Electrochem. Commun., 2008, ?10(2): 293-297
37 DENG Li-Fang, LI Fang-Bai, ZHOU Shun-Gui, HUANG De-Yin, NI Jin-Ren. Chinese Science Bulletin, 2009, ?(19): 2983-2987
邓丽芳, 李芳柏, 周顺桂, 黄德银, 倪晋仁. 科学通报, 2009, ?(19): 2983-2987
38 Call D F, Logan B E. Biosensors & Bioelectronics, 2011, ?26(11): 4526-4531
39 Holmes D E, Nicoll J S, Bond D R, Lovley D R. Appl. Environ. Microbiol., 2004, ?70(10): 6023-6030
40 Holmes D E, Bond D R, Lovley D R. Appl. Environ. Microbiol., 2004, ?70(2): 1234-1237
41 Fedorovich V, Knighton M C, Pagaling E, Ward F B, Free A, Goryanin I. Appl. Environ. Microbiol., 2009, ?75(23): 7326-7334
42 Niessen J, Schroder U, Scholz F. Electrochem. Commun., 2004, ?6(9): 955-958
43 Bond D R, Lovley D R. Appl. Environ. Microbiol., 2005, ?71(4): 2186-2189
44 Sayed E T, Tsujiguchi T, Nakagawa N. Bioelectrochem., 2012, ?86: 97-101
45 Kim B H, Kim H J, Hyun M S, Park D H. J. Microbiol. Biotechnol., ?1999, ?9(2): 127-131
46 Liu Y, Harnisch F, Fricke K, Sietmann R, Schroder U. Biosens. Bioelectron., 2008, ?24(4): 1006-1011
47 Niessen J, Schroder U, Harnisch F, Scholz F. Letters in Applied Microbiology, 2005, ?41(3): 286-290
48 Ren Z, Steinberg L M, Regan J M. Water Science and Technology, 2008, ?58(3): 617-622
49 Marsili E, Baron D B, Shikhare I D, Coursolle D, Gralnick J A, Bond D R. PNAS, 2008, ?105(10): 3968-3973
50 Kim H J, Park H S, Hyun M S, Chang I S, Kim M, Kim B H. Enzyme and Microbial Technology, 2002, ?30(2): 145-152
51 Rabaey K, Boon N, Siciliano S D, Verhaege M, Verstraete W. Appl. Environ. Microbiol., 2004, ?70(9): 5373-5382
52 Pham C A, Jung S J, Phung N T, Lee J, Chang I S, Kim B H, Yi H, Chun J. FEMS Microbiol. Lett., 2003, ?223(1): 129-134
53 Gomelsky M, Kaplan S. Journal of Bacteriology, ?1996, ?178(7): 1946-1954
54 ZHANG Ling-Hua, KUANG Zhe-Shi, CHEN Wei, ZHANG Bao-Ling. Journal of South China Normal University, ?2001, ?(4): 37-39
张玲华, 邝哲师, 陈 薇, ?张宝玲. 华南师范大学学报 (自然科学版) , 2001, ?4: 37-39
55 Gumaelius L, Magnusson G, Pettersson B, Dalhammar G. International Journal of Systematic and Evolutionary Microbiology, 2001, ?51(3): 999-1006
56 Lee S A, Choi Y, Jung S H, Kim S. Bioelectrochem., 2002, ?57(2): 173-178
57 Walker A L, Walker Jr C W. Journal of Power Sources, 2006, ?160(1): 123-129
58 YIN Yun, LIU Yi-Sheng, WANG Yi-Fei, LI Wei, ZHENG Xiao-Dong. Chinese Journal of Applied & Environmental Biology, 2010, ?16(3): 412-414
殷 赟, 刘宜胜, 王一非, 李 伟, 郑晓冬. 应用与环境生物学报, 2010, ?16, (3): 412-414
59 Prasad D, Arun S, Murugesan M, Padmanaban S, Satyanarayanan R, Berchmans S, Yegnaraman V. Biosensors and Bioelectronics, 2007, ?22(11): 2604-2610
60 Touch N, Hibino T, Nagatsu Y, Tachiuchi K. Bioresource Technology, 2014, ?158: 225-230
61 Bond D R, Holmes D E, Tender L M, Lovley D R. Science, 2002, ?295(5554): 483-485
62 Cheng S, Logan B E. PNAS, 2007, ?104(47): 18871-18873
63 Moon H, Chang I S, Jang J K, Kim K S, Lee J, Lovitt R W, Kim B H. J. Microbiol. Biotechnol., 2005, ?15(1): 192-196
64 Villano M, Aulenta F, Ciucci C, Ferri T, Giuliano A, Majone M. Bioresource Technology, 2010, 101(9): 3085-3090〖ZK)〗
Application of Electrochemically Active Bacteria as
Anodic Biocatalyst in Microbial Fuel Cells
ZHANG Yi-Chi, JIANG Zhao-Hong, LIU Ying*
(College of Life Sciences, Northwest A & F University, Yangling 711100, China)
Abstract Microbial fuel cell (MFC) is a novel device with the function to produce energy and degrade organic materials. The characteristics of anodic electrochemically active bacteria and catalytic activity are one of key factors to affect MFC performance. This review summarized the enrichment, source, taxonomy, physiological and biochemical characteristics and electricity production ability of electrochemically active bacteria.
Keywords Microbial fuel cell; Electrochemically active bacteria; Dissimilatory iron-reducing bacteria; Anode respiring bacteria; Review
(Received 19 July 2014; accepted 7 October 2014)
This work was supported by the National Natural Science Foundation of China (No.21375107)
