无刺构骨种子的休眠与萌发研究
况红玲 杜丹 张欣 喻峰 王昆 杨玉洁 胡津鑫


摘 要: 冬青科植物种子在自然条件下,需要隔年甚至3年才能实现萌发,本文以无刺构骨种子为研究对象,通过对无刺构骨种子种皮的透性(透气性和透水性)、不同时间的酸蚀处理、不同浓度的GA3溶液以及层积处理等多方面的试验,寻找阻碍种子萌发的因素,了解无刺构骨种子的休眠原因,初步了解无刺构骨种子的萌发生理。试验结果表明:无刺构骨种子经先暖温后低温及层积处理更有利于种子的生理后熟和萌发。
关键词: 无刺构骨;种子萌发;酸蚀处理;层积处理;GA3处理
中图分类号:S792 文献标识码:A 文章编号:1004-3020(2018)01-0014-05
Study on Dormancy and Germination of Seeds with Ilex cornuta var.fortunei
Kuang Hongling(1) Du Dan(1) Zhang Xin(1) Yu Feng(1) Wang Kun(1) Yang Yujie(2) Hu Jinxin(2)
(1. Faye Ecological Environment Group Co., LTD Wuhan 430206;2. Yangtze University Jingzhou 434100)
Abstract: The seed of Pinus elliottii seeds under the natural conditions is necessary for the next year or even three years to achieve germination. In this paper, the seeds of the seeds without acupuncture are used to study the permeability (permeability and permeability) Time agglutination treatment, different concentrations of GA3 solution and chromatography treatment and other aspects of the experiment to find the factors that impede the seed germination, to understand the seeds of the seeds without acupuncture, and to understand the germination and physiology of seeds without acupuncture. The results showed that the low temperature and stratification treatment of the seeds without the first warm temperature was more favorable to the physiological maturation and germination of the seeds.
Key words: Ilex cornuta var. fortunei;seed germination;acid etching treatment;lamination treatment; GA3 treatment
無刺枸骨(Ilex cornuta var.fortunei)是冬青科(Aquifoliaceae)冬青属枸骨(I. cornuta)的自然变种,常绿灌木或小乔木,树种枝繁叶茂,叶形奇特,树叶浓绿有光泽。树冠圆整,4~5月开黄绿色小花,花序为聚伞花序或伞形花序,雌雄异株,核果球形,初为绿色,入秋成熟转红,满枝累累硕果,鲜艳夺目,经冬不凋直至来年春季,经修枝整形可制作成大树形,球形及树状盆景,是良好的观花、观果、观叶、观形的树种,多应用于庭园观赏和城市绿化。
由于无刺枸骨种子的休眠周期一般长达3年,极大影响了生产上的繁殖效率。如何快速打破种子休眠,提高种子的发芽率,是目前无刺枸骨播种繁殖研究的难点。本文通过对无刺构骨种子的深入了解,探究影响种子休眠的因素,寻找快速打破种子休眠的合理方法,同时为冬青属其它树种种子如何打破休眠奠定基础。
1 材料与方法
1.1 材料
供使用的无刺构骨种子为2016年11月从市场购入的当年新鲜种子,核果用水浸渍2~3 d,待果肉充分软熟后揉搓,淘洗,去除上层漂浮种子,沉在下层的饱满种子为试验材料。
1.2 方法
(1)种子千粒重、形态及生活力的测定。
种子的千粒重及形态测定:使用种子百粒法来测定,以100粒为一组,设五个重复。检测并记录种子的长宽,种壳重以及胚乳的大小。
种子生活力的测定:随机取2×100粒精选种子,浸水48 h,横向切去种子顶端2/3~3/4;而后将种子置于35 ℃无光培养箱中,02%四唑溶液染色12 h;染色结束后流水反复冲洗,剥除种皮,沿中轴线纵切种子,露出胚等组织,在体视显微镜下观察,并拍照。鉴定标准为:
①有生活力种子:胚和胚乳全部正常染色;胚全部染色,胚乳少量未染色。
②无生活力种子:胚和胚乳都未染色;胚全部染色,胚乳未染色。
(2)种子吸水性的测定。
分别取风干种子两份,60粒为一份,第一份为完整的风干种子,第二份用针刺破种皮,将两份种子置于25 ℃恒温培养箱中吸水,分别在2,4,6,8,10,12 h)测重。取出种子时,吸水纸彻底吸干种子表面的浮水后静置10 min称重,并做好记录。
(3)浓硫酸处理。
将供试验的种子倒入一定量的浓硫酸(比重1.84)溶液中,经过不同时间(1 min,5 min,10 min,30 min,1 h,2 h,3 h)处理后种子再置于流水冲洗16~20 h,再进行发芽试验,该处理以20粒为一组,三次重复。
(4)药剂处理。
将供试验的冬青种子用GA3溶液处理,经过不同浓度的GA3+H2SO4溶液(100,250,500 mg/L)浸种24 h,再行发芽试验,该处理以20粒为一组,三次重复。
(5)层积处理。
自然层级:1月1日, 将种子与湿沙分层堆放(湿沙与种子的比例为3∶1)于花盆内,盆口用塑料布扎紧,置于室外干燥避风阴凉处、排水良好、深20 cm的土坑中催芽约180 d,7月1日取出种子统计发芽率,该处理以100粒为一组,三次重复。
低温层级:1月1日,将种子与湿沙分层堆放(湿沙与种子的比例为3∶1)于花盆内,盆口用塑料布扎紧,置于室内冰箱内,催芽180 d,温度为4 ℃,7月1日取出,统计发芽率,该处理以100粒为一组,三次重复。
變温层积:1月1日,将种子与湿沙分层堆放(湿沙与种子的比例为3∶1)于花盆内,盆口用塑料布扎紧,置于室内冰箱内,催芽120 d,温度为4 ℃,5月1日从冰箱中取出,置于室外干燥避风阴凉处、排水良好、深20 cm的土坑中催芽60 d,统计发芽率,该处理以100粒为一组,三次重复。
1.3 数据统计分析
数据采用Excel、SAS软件进行统计和分析。
萌发率(%)=发芽种子数/测定种子数×100%
吸水率(%)=[吸水后质量(g)-吸水前质量(g)]/吸水前质量(g)×100%
2 结果与分析
2.1 无刺构骨种子生物学特性
(1)无刺构骨种子的基本形态特征。无刺构骨果实为浆果状,通常为球形,成熟的红果悬挂于枝头,干后变为深红色,果肉肥厚多汁,每颗果实大约有4~5粒种子,种子狭长披针形,外背面形如沟壑。
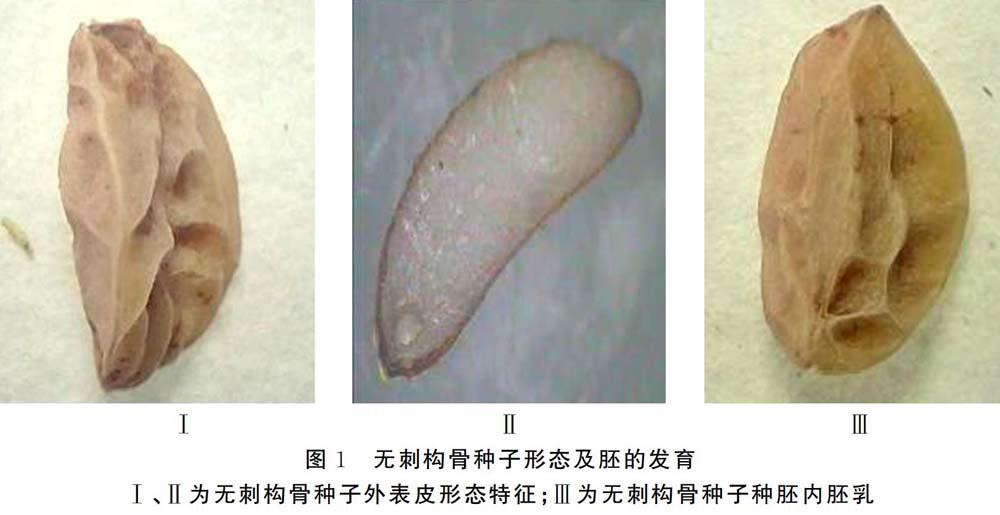
通过对无刺构骨果实的揉搓和清洗,得到干净良好的种子,通过对种子的长度、宽度以及胚乳的测定和观察,得到以下数据,见图1。
种子外部形态统一,长度(664±035)mm,宽度(386±03)mm,高度(372±025)mm,千粒重为1522 g,种子有仁率7826%,种壳的重量占完整无刺构骨种子的81%,结果表明,种壳的厚重在一定程度上导致了种子的休眠。
(2)种子生活力测定。种子生活力是检验种子生长发育好坏的基本标准,同时也反应种子的质量问题,经测试,用于试验的种子中有生活力的为187粒,无生活力的为13粒,有生活力的种子占到总种子数量的93.5%,说明供试验的大多数种子的质量良好。
(3)种子的透水性。完整种子和破皮种子的吸水情况,见图2。
由图2所示,在浸泡前期,完整枸骨种子与破皮种子的吸水情况基本保持一致,随着时间的推移,在浸泡中期,破皮的无刺构骨种子的吸水率高于完整种子,在浸泡后期,两种种子吸水率达到饱和一致。结果表明:由于无刺构骨种子种皮坚硬,完整种子的吸水率增长缓慢,种皮的坚硬程度在一定程度上对种子的吸水率有影响。
2.2 不同酸蚀时间处理对种子发芽率的影响
由于无刺构骨种子种皮木质而坚硬,阻碍种子的萌发,但酸蚀处理能够使种壳减轻,增加它的透性,提高种子的发芽率。通过对无刺构骨种子不同时间的酸蚀处理,可摸索出最合适的酸蚀时间,见下表1。
据表1所知,无刺构骨种子在经过酸蚀处理后,酸蚀时间越久,种壳的腐蚀比率越多,腐烂率也相对增加,而发芽率随着种壳腐蚀比率和腐烂率的变化而变化,试验结果表明:无刺构骨种子经过酸蚀处理时间1 h,更合适地减轻种壳的重量,稳定种子的腐烂程度,促进无刺构骨种子的萌发。
2.3 GA3和浓硫酸的综合处理对种子发芽率的影响
据表2可知,不同浓度GA3溶液与不同酸蚀时间的综合处理对无刺构骨种子发芽率有着明显的提升,发芽率随着GA3浓度的增加而提高,但不是浓度越高越有利,浓度太高反而降低了无刺构骨种子胚的活性,结果表明:用浓度为250 ml GA3处理的种子萌发效果最好。
2.4 层积处理对种子发芽率的影响
层积处理是打破种子综合性休眠的有效方法,层积处理可分为自然层积、低温层积和变温层积,通过对不同的层积处理(自然层积、低温层积180 d,变温层积暖温60 d+低温120 d)探讨对无刺构骨种子发芽率的影响,如图3所示。
实验表明:无刺构骨种子经过自然层积、低温层积180 d后,种子的发芽率分别为06%、13%,相比较自然层积与低温层积,变温层积的萌发率明显有优势,发芽率达5.8%,于是可推断无刺构骨种子经过自然层积,对无刺构骨种子打破休眠无明显的促进作用,与自然层积处理相比,低温层积可使部分种子打破休眠,但发芽率也是较低,然而,经过变温层积(先暖温再低温)层积处理后的无刺构骨种子,更有利于种子胚的生长发育,提高种子的发芽率。
3 讨论与结论
3.1 讨论
(1)种皮对种子休眠的影响。由于无刺构种子种皮木质化,坚硬而紧密等一系列的种皮障碍,导致种子进行强迫休眠,综合而言,种皮障碍通常指的是种皮的不透水性、不透气性和种皮产生的机械障碍所引起的。坚硬的种皮限制了种胚与外界的气体交换,阻断氧气和二氧化碳的的排放,阻碍种胚的呼吸,无法为种子萌发提供必需的能量物质,很多学者表明,种子坚硬木质化、细胞排列致密的种皮特性是导致种子休眠的主要因素,根据同对李近雨对核桃种子的研究表明,种壳的机械约束是种子发芽慢的主要原因[6],狭叶冬青(Ilex fargesii )坚硬致密的种皮透气性较差是种子强迫休眠的因素之一[7],徐本美等的试验指出大果种子的强迫休眠主要是种皮透气性差所致,此特性与山楂(Crata- egus pinnatifida bunge.)、圆柏(Sabina chinensis)、蔷薇(Rosa laevigata)、秤锤树(S.xylocarpa)、大叶冬青、膀胱果(S.holocarpa)相近[8-10]等学说进行分析和比较,本试验中的无刺构骨种子存在的不透水性、不透气性和种皮的机械障碍在很大程度上阻碍种子无法萌发,抑制胚的生长。
酸蝕处理能够有效的酸蚀种皮外致密组织细胞,减少种皮障碍和种壳的重量,使胚能够脱离种皮的束缚,增强胚的透性,促进种子胚的萌发[11],对无刺构骨种子经过浓硫酸处理后,种子的发芽率得到大大地提高,并且把握合适的酸蚀时间是至关重要的,若酸蚀时间不够,则无法充分的酸蚀种皮障碍,为种子萌发达到预期的效果,若酸蚀时间太久,则会充分酸蚀掉种皮,对种胚造成不可挽回的损害,容易使种胚腐烂,导致种胚死亡。
(2)胚发育状况对种子休眠的影响。在无刺构骨果实成熟后进行采摘,揉搓果实,风干种子,以便解剖种子,如图1所示,种胚长度约0.3~0.4mm,仅为萌发时胚长的1/10,此时胚尚处于胚原基阶段;幼胚从胚乳的营养物质中摄取能量,以促进种子萌发,种子在自然成熟后,种胚还在初级阶段,胚未发育完全,此时需要先完成形态发育成熟才能打破休眠而促进萌发。
对于如何加快无刺构骨种子胚生理后熟的过程,变温层积处理是最有效的处理方法。暖层积阶段时,种胚才开始慢慢地分化,种胚生长发育较慢,这与Leonard 等[12]进行西洋参种子研究中所获结论一致。暖层积过程中胚可能会出现一段时间的静止期,因此暖层积初期胚乳需要从种胚汲取大量的营养,于是种胚生长缓慢。低温层积阶段种胚开始迅速生长,层积120 d后,大部分的无刺构骨种子已经完成生理后熟的过程,但不同的是,各种胚的质量和胚的发育状况有所不同,小部分无刺构骨种子仍出现半成熟状态,结果导致无刺构骨种子萌发齐性不一致。
(3)内源抑制物对种子休眠的影响。许多植物的果实和种皮中都存在发芽抑制物,主要包含一些有机酸、生物碱、脂类、醛类等物质具有直接或者间接抑制种子萌发的作用[13],其中香豆素、脱落酸是两种对种子萌发较强的抑制物[14]。
GA3处理能够有效的促进种子胚的生长发育,它能诱导胚乳产生水解酶,将水中的贮藏物质从大分子转化为小分子,如淀粉水解为糖,蛋白质水解为氨基酸,来供胚进行吸收,同时促进胚中有机物和无机物间的转换,使胚完成生理后熟过程,还能促进细胞分裂和伸长,促进DNA和RNA淀粉合成[15-16],能够抑制ABA在休眠种子中的积累[17]。
3.2 结论
通过对无刺构骨种子一系列的探讨和研究,来找寻影响种子休眠的主要因素,无刺构骨种子具有种皮坚硬的特性,种壳占种子重量的87.3%,透气性极差,影响种子胚和胚乳的物质和气体交换,阻碍种子的萌发,是导致种子休眠的主要因素之一。酸蚀处理能够酸蚀种皮,打破种皮障碍,加快种胚的生长发育,但并不能使种子提前萌发,种子的胚和胚乳中存在一定的内源抑制物,在一定程度上限制胚的生长发育,赤霉素GA3能够减少胚中的内源抑制物,促进胚的生长发育,另外,因为无刺构骨种子具有隔年发芽的特性,采摘后的果实存在未完全发育的情况,经过生理后熟才能进行萌发,层积处理是一种常用的打破种子休眠的方法,它分为自然层积、低温层积和变温层积,由于,无刺构骨种子具有隔年发芽生理特性和生理后熟的种皮特性,自然层积不能短时间内的打破种子休眠,低温层积只能打破部分成熟种子的休眠,且发芽率低,变温层积(先暖温后低温)处理对打破种子休眠有非常显著的变化,能够加快种子生长发育,加快种子胚的吸收,促进种子萌发,是最适合作为打破种子休眠的一种层积处理。因此,在了解阻碍无刺构骨种子休眠的种种因素之后,得出最佳的解除种子休眠的方法为:98%浓硫酸1 h+250 mg/L赤霉素溶液浸种24 h+暖层积60 d+低温层积120 d。
参 考 文 献
[1]刘祖生,梁月荣,徐月荣,等.苦丁茶化学成分的研究——游离氨基酸分析[J].浙江农业大学学报,1998:71-74.
[2]曾沧江.中国冬青科植物志资料[J].植物研究,1981(Z):1-21.
[3]郁建平.贵州苦丁茶植物资源及化学成分分析[J].植物资源与环境,1997(3):23-26.
[4]戴素贤,谢赤军,袁学培,等.苦丁茶香气的化学组分[J].植物资源与环境,1998(1):68-73.
[5]周晓峰.几种冬青属树种种子休眠原因及萌发特性研究[J].南京林业大学,2010(6):17-18.
[6]李近雨.核桃种子烂种原因及发芽条件的研究[J].林业科学,1994,30(1):18-24.
[7]何彦峰.狭叶冬青种子休眠与萌发的研究[J].浙江林业科技,2008,28(3):63-65.
[8]徐本美,史晓华,黎念林.大叶冬青种子的休眠与萌发初探[J].植物引种驯化集刊,1997(11):150.
[9]徐本美,张治明,白克智,等.关于木本植物种子休眠与萌发的研究[M].植物引种驯化集刊(第十集),北京:科学出版社,1995:123-129.
[10]徐本美,史晓华,孙运涛.膀胱果种子休眠与萌发的研究[J].种子,2002(1):13-14.
[11]范川,李贤伟.珙桐的研究现状及展望[J].林业科技,2004,29(6):55-58.
[12]李正理,李荣傲.黄连种子后熟过程的解剖学研究[J].植物学报,1985,27(2):122-127.
[13]余朝霞,黄雪群,方志尚,等.GA3对林木种子萌发的调控研究进展[J].浙江林业科技,2003,23(1):73-76.
[14]傅强,杨期和,叶万辉.种子休眠的解除方法[J].广西农业生物科学,2003,22(3):230-234.
[15]AlexiosA. Alexopoulos, George Aivalakis, Konstantions. A. Akoumianakis,et.al.Effect of gobberellic acid on the duration of dormancy of potato tubers produced by plants from true potato seed[J].Postharvest Biology and Technology,2008,49:424-430.
[16]Maarten Koornneef, Leonie Bentstink Henk Hilhorst. Seed Dormancy and germination [J].Current Opinion in Plant Biology, 2002,5:33-36.
[17]Bent Sink L, Hilhorst H, Seed dormancy and germination[J],Current Opinion in Plant Biology,2002,5:33-36.
摘 要: 冬青科植物种子在自然条件下,需要隔年甚至3年才能实现萌发,本文以无刺构骨种子为研究对象,通过对无刺构骨种子种皮的透性(透气性和透水性)、不同时间的酸蚀处理、不同浓度的GA3溶液以及层积处理等多方面的试验,寻找阻碍种子萌发的因素,了解无刺构骨种子的休眠原因,初步了解无刺构骨种子的萌发生理。试验结果表明:无刺构骨种子经先暖温后低温及层积处理更有利于种子的生理后熟和萌发。
关键词: 无刺构骨;种子萌发;酸蚀处理;层积处理;GA3处理
中图分类号:S792 文献标识码:A 文章编号:1004-3020(2018)01-0014-05
Study on Dormancy and Germination of Seeds with Ilex cornuta var.fortunei
Kuang Hongling(1) Du Dan(1) Zhang Xin(1) Yu Feng(1) Wang Kun(1) Yang Yujie(2) Hu Jinxin(2)
(1. Faye Ecological Environment Group Co., LTD Wuhan 430206;2. Yangtze University Jingzhou 434100)
Abstract: The seed of Pinus elliottii seeds under the natural conditions is necessary for the next year or even three years to achieve germination. In this paper, the seeds of the seeds without acupuncture are used to study the permeability (permeability and permeability) Time agglutination treatment, different concentrations of GA3 solution and chromatography treatment and other aspects of the experiment to find the factors that impede the seed germination, to understand the seeds of the seeds without acupuncture, and to understand the germination and physiology of seeds without acupuncture. The results showed that the low temperature and stratification treatment of the seeds without the first warm temperature was more favorable to the physiological maturation and germination of the seeds.
Key words: Ilex cornuta var. fortunei;seed germination;acid etching treatment;lamination treatment; GA3 treatment
無刺枸骨(Ilex cornuta var.fortunei)是冬青科(Aquifoliaceae)冬青属枸骨(I. cornuta)的自然变种,常绿灌木或小乔木,树种枝繁叶茂,叶形奇特,树叶浓绿有光泽。树冠圆整,4~5月开黄绿色小花,花序为聚伞花序或伞形花序,雌雄异株,核果球形,初为绿色,入秋成熟转红,满枝累累硕果,鲜艳夺目,经冬不凋直至来年春季,经修枝整形可制作成大树形,球形及树状盆景,是良好的观花、观果、观叶、观形的树种,多应用于庭园观赏和城市绿化。
由于无刺枸骨种子的休眠周期一般长达3年,极大影响了生产上的繁殖效率。如何快速打破种子休眠,提高种子的发芽率,是目前无刺枸骨播种繁殖研究的难点。本文通过对无刺构骨种子的深入了解,探究影响种子休眠的因素,寻找快速打破种子休眠的合理方法,同时为冬青属其它树种种子如何打破休眠奠定基础。
1 材料与方法
1.1 材料
供使用的无刺构骨种子为2016年11月从市场购入的当年新鲜种子,核果用水浸渍2~3 d,待果肉充分软熟后揉搓,淘洗,去除上层漂浮种子,沉在下层的饱满种子为试验材料。
1.2 方法
(1)种子千粒重、形态及生活力的测定。
种子的千粒重及形态测定:使用种子百粒法来测定,以100粒为一组,设五个重复。检测并记录种子的长宽,种壳重以及胚乳的大小。
种子生活力的测定:随机取2×100粒精选种子,浸水48 h,横向切去种子顶端2/3~3/4;而后将种子置于35 ℃无光培养箱中,02%四唑溶液染色12 h;染色结束后流水反复冲洗,剥除种皮,沿中轴线纵切种子,露出胚等组织,在体视显微镜下观察,并拍照。鉴定标准为:
①有生活力种子:胚和胚乳全部正常染色;胚全部染色,胚乳少量未染色。
②无生活力种子:胚和胚乳都未染色;胚全部染色,胚乳未染色。
(2)种子吸水性的测定。
分别取风干种子两份,60粒为一份,第一份为完整的风干种子,第二份用针刺破种皮,将两份种子置于25 ℃恒温培养箱中吸水,分别在2,4,6,8,10,12 h)测重。取出种子时,吸水纸彻底吸干种子表面的浮水后静置10 min称重,并做好记录。
(3)浓硫酸处理。
将供试验的种子倒入一定量的浓硫酸(比重1.84)溶液中,经过不同时间(1 min,5 min,10 min,30 min,1 h,2 h,3 h)处理后种子再置于流水冲洗16~20 h,再进行发芽试验,该处理以20粒为一组,三次重复。
(4)药剂处理。
将供试验的冬青种子用GA3溶液处理,经过不同浓度的GA3+H2SO4溶液(100,250,500 mg/L)浸种24 h,再行发芽试验,该处理以20粒为一组,三次重复。
(5)层积处理。
自然层级:1月1日, 将种子与湿沙分层堆放(湿沙与种子的比例为3∶1)于花盆内,盆口用塑料布扎紧,置于室外干燥避风阴凉处、排水良好、深20 cm的土坑中催芽约180 d,7月1日取出种子统计发芽率,该处理以100粒为一组,三次重复。
低温层级:1月1日,将种子与湿沙分层堆放(湿沙与种子的比例为3∶1)于花盆内,盆口用塑料布扎紧,置于室内冰箱内,催芽180 d,温度为4 ℃,7月1日取出,统计发芽率,该处理以100粒为一组,三次重复。
變温层积:1月1日,将种子与湿沙分层堆放(湿沙与种子的比例为3∶1)于花盆内,盆口用塑料布扎紧,置于室内冰箱内,催芽120 d,温度为4 ℃,5月1日从冰箱中取出,置于室外干燥避风阴凉处、排水良好、深20 cm的土坑中催芽60 d,统计发芽率,该处理以100粒为一组,三次重复。
1.3 数据统计分析
数据采用Excel、SAS软件进行统计和分析。
萌发率(%)=发芽种子数/测定种子数×100%
吸水率(%)=[吸水后质量(g)-吸水前质量(g)]/吸水前质量(g)×100%
2 结果与分析
2.1 无刺构骨种子生物学特性
(1)无刺构骨种子的基本形态特征。无刺构骨果实为浆果状,通常为球形,成熟的红果悬挂于枝头,干后变为深红色,果肉肥厚多汁,每颗果实大约有4~5粒种子,种子狭长披针形,外背面形如沟壑。
通过对无刺构骨果实的揉搓和清洗,得到干净良好的种子,通过对种子的长度、宽度以及胚乳的测定和观察,得到以下数据,见图1。
种子外部形态统一,长度(664±035)mm,宽度(386±03)mm,高度(372±025)mm,千粒重为1522 g,种子有仁率7826%,种壳的重量占完整无刺构骨种子的81%,结果表明,种壳的厚重在一定程度上导致了种子的休眠。
(2)种子生活力测定。种子生活力是检验种子生长发育好坏的基本标准,同时也反应种子的质量问题,经测试,用于试验的种子中有生活力的为187粒,无生活力的为13粒,有生活力的种子占到总种子数量的93.5%,说明供试验的大多数种子的质量良好。
(3)种子的透水性。完整种子和破皮种子的吸水情况,见图2。
由图2所示,在浸泡前期,完整枸骨种子与破皮种子的吸水情况基本保持一致,随着时间的推移,在浸泡中期,破皮的无刺构骨种子的吸水率高于完整种子,在浸泡后期,两种种子吸水率达到饱和一致。结果表明:由于无刺构骨种子种皮坚硬,完整种子的吸水率增长缓慢,种皮的坚硬程度在一定程度上对种子的吸水率有影响。
2.2 不同酸蚀时间处理对种子发芽率的影响
由于无刺构骨种子种皮木质而坚硬,阻碍种子的萌发,但酸蚀处理能够使种壳减轻,增加它的透性,提高种子的发芽率。通过对无刺构骨种子不同时间的酸蚀处理,可摸索出最合适的酸蚀时间,见下表1。
据表1所知,无刺构骨种子在经过酸蚀处理后,酸蚀时间越久,种壳的腐蚀比率越多,腐烂率也相对增加,而发芽率随着种壳腐蚀比率和腐烂率的变化而变化,试验结果表明:无刺构骨种子经过酸蚀处理时间1 h,更合适地减轻种壳的重量,稳定种子的腐烂程度,促进无刺构骨种子的萌发。
2.3 GA3和浓硫酸的综合处理对种子发芽率的影响
据表2可知,不同浓度GA3溶液与不同酸蚀时间的综合处理对无刺构骨种子发芽率有着明显的提升,发芽率随着GA3浓度的增加而提高,但不是浓度越高越有利,浓度太高反而降低了无刺构骨种子胚的活性,结果表明:用浓度为250 ml GA3处理的种子萌发效果最好。
2.4 层积处理对种子发芽率的影响
层积处理是打破种子综合性休眠的有效方法,层积处理可分为自然层积、低温层积和变温层积,通过对不同的层积处理(自然层积、低温层积180 d,变温层积暖温60 d+低温120 d)探讨对无刺构骨种子发芽率的影响,如图3所示。
实验表明:无刺构骨种子经过自然层积、低温层积180 d后,种子的发芽率分别为06%、13%,相比较自然层积与低温层积,变温层积的萌发率明显有优势,发芽率达5.8%,于是可推断无刺构骨种子经过自然层积,对无刺构骨种子打破休眠无明显的促进作用,与自然层积处理相比,低温层积可使部分种子打破休眠,但发芽率也是较低,然而,经过变温层积(先暖温再低温)层积处理后的无刺构骨种子,更有利于种子胚的生长发育,提高种子的发芽率。
3 讨论与结论
3.1 讨论
(1)种皮对种子休眠的影响。由于无刺构种子种皮木质化,坚硬而紧密等一系列的种皮障碍,导致种子进行强迫休眠,综合而言,种皮障碍通常指的是种皮的不透水性、不透气性和种皮产生的机械障碍所引起的。坚硬的种皮限制了种胚与外界的气体交换,阻断氧气和二氧化碳的的排放,阻碍种胚的呼吸,无法为种子萌发提供必需的能量物质,很多学者表明,种子坚硬木质化、细胞排列致密的种皮特性是导致种子休眠的主要因素,根据同对李近雨对核桃种子的研究表明,种壳的机械约束是种子发芽慢的主要原因[6],狭叶冬青(Ilex fargesii )坚硬致密的种皮透气性较差是种子强迫休眠的因素之一[7],徐本美等的试验指出大果种子的强迫休眠主要是种皮透气性差所致,此特性与山楂(Crata- egus pinnatifida bunge.)、圆柏(Sabina chinensis)、蔷薇(Rosa laevigata)、秤锤树(S.xylocarpa)、大叶冬青、膀胱果(S.holocarpa)相近[8-10]等学说进行分析和比较,本试验中的无刺构骨种子存在的不透水性、不透气性和种皮的机械障碍在很大程度上阻碍种子无法萌发,抑制胚的生长。
酸蝕处理能够有效的酸蚀种皮外致密组织细胞,减少种皮障碍和种壳的重量,使胚能够脱离种皮的束缚,增强胚的透性,促进种子胚的萌发[11],对无刺构骨种子经过浓硫酸处理后,种子的发芽率得到大大地提高,并且把握合适的酸蚀时间是至关重要的,若酸蚀时间不够,则无法充分的酸蚀种皮障碍,为种子萌发达到预期的效果,若酸蚀时间太久,则会充分酸蚀掉种皮,对种胚造成不可挽回的损害,容易使种胚腐烂,导致种胚死亡。
(2)胚发育状况对种子休眠的影响。在无刺构骨果实成熟后进行采摘,揉搓果实,风干种子,以便解剖种子,如图1所示,种胚长度约0.3~0.4mm,仅为萌发时胚长的1/10,此时胚尚处于胚原基阶段;幼胚从胚乳的营养物质中摄取能量,以促进种子萌发,种子在自然成熟后,种胚还在初级阶段,胚未发育完全,此时需要先完成形态发育成熟才能打破休眠而促进萌发。
对于如何加快无刺构骨种子胚生理后熟的过程,变温层积处理是最有效的处理方法。暖层积阶段时,种胚才开始慢慢地分化,种胚生长发育较慢,这与Leonard 等[12]进行西洋参种子研究中所获结论一致。暖层积过程中胚可能会出现一段时间的静止期,因此暖层积初期胚乳需要从种胚汲取大量的营养,于是种胚生长缓慢。低温层积阶段种胚开始迅速生长,层积120 d后,大部分的无刺构骨种子已经完成生理后熟的过程,但不同的是,各种胚的质量和胚的发育状况有所不同,小部分无刺构骨种子仍出现半成熟状态,结果导致无刺构骨种子萌发齐性不一致。
(3)内源抑制物对种子休眠的影响。许多植物的果实和种皮中都存在发芽抑制物,主要包含一些有机酸、生物碱、脂类、醛类等物质具有直接或者间接抑制种子萌发的作用[13],其中香豆素、脱落酸是两种对种子萌发较强的抑制物[14]。
GA3处理能够有效的促进种子胚的生长发育,它能诱导胚乳产生水解酶,将水中的贮藏物质从大分子转化为小分子,如淀粉水解为糖,蛋白质水解为氨基酸,来供胚进行吸收,同时促进胚中有机物和无机物间的转换,使胚完成生理后熟过程,还能促进细胞分裂和伸长,促进DNA和RNA淀粉合成[15-16],能够抑制ABA在休眠种子中的积累[17]。
3.2 结论
通过对无刺构骨种子一系列的探讨和研究,来找寻影响种子休眠的主要因素,无刺构骨种子具有种皮坚硬的特性,种壳占种子重量的87.3%,透气性极差,影响种子胚和胚乳的物质和气体交换,阻碍种子的萌发,是导致种子休眠的主要因素之一。酸蚀处理能够酸蚀种皮,打破种皮障碍,加快种胚的生长发育,但并不能使种子提前萌发,种子的胚和胚乳中存在一定的内源抑制物,在一定程度上限制胚的生长发育,赤霉素GA3能够减少胚中的内源抑制物,促进胚的生长发育,另外,因为无刺构骨种子具有隔年发芽的特性,采摘后的果实存在未完全发育的情况,经过生理后熟才能进行萌发,层积处理是一种常用的打破种子休眠的方法,它分为自然层积、低温层积和变温层积,由于,无刺构骨种子具有隔年发芽生理特性和生理后熟的种皮特性,自然层积不能短时间内的打破种子休眠,低温层积只能打破部分成熟种子的休眠,且发芽率低,变温层积(先暖温后低温)处理对打破种子休眠有非常显著的变化,能够加快种子生长发育,加快种子胚的吸收,促进种子萌发,是最适合作为打破种子休眠的一种层积处理。因此,在了解阻碍无刺构骨种子休眠的种种因素之后,得出最佳的解除种子休眠的方法为:98%浓硫酸1 h+250 mg/L赤霉素溶液浸种24 h+暖层积60 d+低温层积120 d。
参 考 文 献
[1]刘祖生,梁月荣,徐月荣,等.苦丁茶化学成分的研究——游离氨基酸分析[J].浙江农业大学学报,1998:71-74.
[2]曾沧江.中国冬青科植物志资料[J].植物研究,1981(Z):1-21.
[3]郁建平.贵州苦丁茶植物资源及化学成分分析[J].植物资源与环境,1997(3):23-26.
[4]戴素贤,谢赤军,袁学培,等.苦丁茶香气的化学组分[J].植物资源与环境,1998(1):68-73.
[5]周晓峰.几种冬青属树种种子休眠原因及萌发特性研究[J].南京林业大学,2010(6):17-18.
[6]李近雨.核桃种子烂种原因及发芽条件的研究[J].林业科学,1994,30(1):18-24.
[7]何彦峰.狭叶冬青种子休眠与萌发的研究[J].浙江林业科技,2008,28(3):63-65.
[8]徐本美,史晓华,黎念林.大叶冬青种子的休眠与萌发初探[J].植物引种驯化集刊,1997(11):150.
[9]徐本美,张治明,白克智,等.关于木本植物种子休眠与萌发的研究[M].植物引种驯化集刊(第十集),北京:科学出版社,1995:123-129.
[10]徐本美,史晓华,孙运涛.膀胱果种子休眠与萌发的研究[J].种子,2002(1):13-14.
[11]范川,李贤伟.珙桐的研究现状及展望[J].林业科技,2004,29(6):55-58.
[12]李正理,李荣傲.黄连种子后熟过程的解剖学研究[J].植物学报,1985,27(2):122-127.
[13]余朝霞,黄雪群,方志尚,等.GA3对林木种子萌发的调控研究进展[J].浙江林业科技,2003,23(1):73-76.
[14]傅强,杨期和,叶万辉.种子休眠的解除方法[J].广西农业生物科学,2003,22(3):230-234.
[15]AlexiosA. Alexopoulos, George Aivalakis, Konstantions. A. Akoumianakis,et.al.Effect of gobberellic acid on the duration of dormancy of potato tubers produced by plants from true potato seed[J].Postharvest Biology and Technology,2008,49:424-430.
[16]Maarten Koornneef, Leonie Bentstink Henk Hilhorst. Seed Dormancy and germination [J].Current Opinion in Plant Biology, 2002,5:33-36.
[17]Bent Sink L, Hilhorst H, Seed dormancy and germination[J],Current Opinion in Plant Biology,2002,5:33-36.
