N-酰基高丝氨酸内酯酶的生物信息学分析
杨艳北 许晶 沈城辉 许继国 饶友生
摘要:采用生物信息学分析方法,对沼泽红假单胞菌N-酰基高丝氨酸内酯酶理化性质和结构特征进行预测。结果表明,该蛋白为稳定的亲水性蛋白,定位在细菌的细胞质中,无信号肽结构。二级结构中含有α-螺旋、β-转角、延伸链和无规则卷曲等结构元件,α-螺旋和无规则卷曲对三级结构的稳定和功能发挥具有重要意义。含有6个磷酸化位点,无跨膜结构域。参与细菌群体感应淬灭。
关键词:沼泽红假单胞菌;N-酰基高丝氨酸内酯酶;生物信息学
中图分类号:Q939.96? ? ?文献标志码:A? ? ?文章编号:1001-1463(2021)02-0031-07
doi:10.3969/j.issn.1001-1463.2021.02.008
Bioinformatics Analysis of N-acyl Homoserine Lactonase
YANG Yanbei 1,2, XU Jing 3, SHEN Chenghui 3, XU Jiguo 1,2, RAO Yousheng 1,2
(1. Institute of Biotechnology Research, Nanchang Normal University, Nanchang Jiangxi 330000, China;? ? ? ? 2. Province Key Lab of Genetic Improvement of Indigenous Chicken Breeds, Nanchang Jiangxi 330000, China;3. Department of Biology, Nanchang Normal University, Nanchang Jiangxi 330000, China)
Abstract:The aim of the present study was to understand the physical and chemical properties and structural characteristic of N-acyl homoserine lactonase in Rhodopseudomonas palustris by bioinformatics method. The results showed that the protein was a stable hydrophilic protein. The protein was localized in the cytoplasm of bacteria. The protein had no signal peptide structure. The secondary structure of the protein contained α-helix, β - angle, extended chain and random coil. The α-helix and random coil played an important role in the stability and function of tertiary structure. The protein contained six phosphorylation sites. The protein had no transmembrane domain. The protein was involved in quorum sensing quenching.
Key words:Rhodopseudomonas palustris;N-acyl homoserine lactonase;Bioinformatics;Biofilm;Analysis
沼澤红假单胞菌可作为微生物饲料添加剂,属于益生菌,广泛地应用于水产养殖 业[1 - 2 ]。生物被膜能够增强益生菌对温度、胃酸和机械剪切力的抵抗能力。生物被膜包埋到微胶囊中创造了新一代的益生菌,国内外学者称之为“第四代益生菌”。微胶囊技术和生物被膜的结合,进一步提高了益生菌对于恶劣环境的抵抗能力。“第四代益生菌”具有更好的抗冷冻干燥能力和耐热性,对于低温储藏和胃液处理,存活率更高[3 ]。群体感应是细菌通过分泌信号分子来调控其不同生物学功能的一种群体行为,例如细菌生物被膜的形成[4 ]。革兰氏阴性细菌群体感应系统主要由N-酰基高丝氨酸内酯类信号分子介导,信号分子的浓度随细菌密度的增加而增加,当达到或超过阈值时,信号分子同受体结合从而触发信号转导,导致细菌生物被膜的形成[5 ]。N-酰基高丝氨酸内酯类信号分子对革兰氏阴性细菌群体感应系统的调控具有十分重要的作用,通过破坏或干扰N-酰基高丝氨酸内酯的合成,达到淬灭群体感应系统的目标。群体感应淬灭通过抑制信号分子的合成,降低信号分子的浓度,降解或修饰信号分子结构,从而破坏细菌群体感应系统的功能。N-酰基高丝氨酸内酯酶作为广泛存在的群体感应淬灭酶,表现出了极强的淬灭活性[6 - 7 ] 。我们选取N-酰基高丝氨酸内酯酶,采用生物信息学的分析方法,对其理化性质、亲水性/疏水性、亚细胞定位、信号肽、二级结构、三级结构、磷酸化位点、跨膜结构域和功能结构域进行分析和预测,旨在为后续研究沼泽红假单胞菌生物被膜形成机制奠定基础,也为生物被膜增强剂的开发提供参考。
1? ?材料与方法
1.1? ?数据来源
沼泽红假单胞菌N-酰基高丝氨酸内酯酶蛋白质序列来自美国国家生物信息中心(NCBI)中已登录的序列,蛋白质序列编码为WP_119858436.1[N-acyl homoserine lactonase family protein (Rhodopseudomonas palustris)]。
1.2? ?研究方法
在线生物信息学预测工具分析N-酰基高丝氨酸内酯酶的蛋白质序列:利用ProtParam(https://web.expasy.org/protparam/)分析理化性质,利用ProtScale (https://web.expasy.org/protscale/)进行亲水性、疏水性的分析和预测,利用PSORTb version 3.0.2 (https://www.psort.org/psortb/)和LocTree3 (https:// www.rostlab.org/services/loctree3/)进行细菌亚细胞定位分析和预测,利用SignalP 4.1 Server(http://www.cbs.dtu.dk/services/SignalP-4.1/)進行信号肽分析和预测,利用SOPMA(https:
//npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl? page=/NPSA/npsa_sopma.html)进行蛋白质二级结构分析和预测,利用SWISS-MODEL (https: //swissmodel. expasy. org/)进行蛋白质三级结构分析和预测,利用NetPhosBac 1.0 server(https://services.healthtech.dtu.dk/service. php?NetPhosBac-1.0)进行细菌蛋白质的磷酸化位点分析和预测。利用TMHMM 2.0 server(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)进行跨膜结构域的分析和预测,利用NCBI的保守域数据库(CDD)(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)进行功能结构域分析和预测。
2? ?结果与分析
2.1? ?N-酰基高丝氨酸内酯酶理化性质分析
利用ProtParam分析了N-酰基高丝氨酸内酯酶的理化性质。N-酰基高丝氨酸内酯酶的分子量为31 488.50 Da,理论等电点为5.14。氨基酸数量为284个,其中含量较多的是甘氨酸(28个,9.9%)、丙氨酸(27,9.5%)、天冬氨酸(24个,8.5%)、亮氨酸(24个,8.5%)。N-酰基高丝氨酸内酯酶含有3个半胱氨酸,推测N-酰基高丝氨酸内酯酶可能具有二硫键结构。N-酰基高丝氨酸内酯酶不具有吡咯赖氨酸和硒半胱氨酸等稀有氨基酸。N-酰基高丝氨酸内酯酶的氨基酸组成分别为丙氨酸(27个,9.5%)、精氨酸(19个,6.7%)、天冬酰胺(7个,2.5%)、天冬氨酸(24个,8.5%)、半胱氨酸(3个,1.1%)、谷氨酰胺(9个,3.2%)、谷氨酸(13个,4.6%)、甘氨酸(28个,9.9%)、组氨酸(8个,2.8%)、异亮氨酸(15个,5.3%)、亮氨酸(24个,8.5%)、赖氨酸(7个,2.5%)、蛋氨酸(8个,2.8%)、苯丙氨酸(15个,5.3%)、脯氨酸(12个,4.2%)、丝氨酸(17个,6.0%)、苏氨酸(15个,5.3%)、色氨酸(7个,2.5%)、酪氨酸(9个,3.2%)、缬氨酸(17个,6.0%)。N-酰基高丝氨酸内酯酶带负电荷氨基酸残基数(天冬酰胺+谷氨酸)为37个,带正电荷氨基酸残基数(精氨酸+赖氨酸)为26个,其分子式为C1407H2143N387O416S11,原子总数为4 364。假设N-酰基高丝氨酸内酯酶所有的胱氨酸残基以半胱氨酸的形式出现,即形成二硫键,N-酰基高丝氨酸内酯酶的消光系数为52 035 L/(mol·cm),吸光度(A280)为1.653;假设二硫键全部打开,N-酰基高丝氨酸内酯酶的消光系数为51 910 L/(mol·cm),吸光度(A280)为1.649。当N-酰基高丝氨酸内酯酶的N端为甲硫氨酸时,在哺乳动物网织红细胞中的半衰期为30 h,在酵母体内的半衰期大于20 h,在大肠杆菌体内半衰期大于10 h。N-酰基高丝氨酸内酯酶的不稳定系数为24.62,小于阈值40,在溶液中性质稳定,推测N-酰基高丝氨酸内酯酶为稳定蛋白。N-酰基高丝氨酸内酯酶脂肪指数为80.42,亲水性平均值为-0.187。亲水性平均值为负值,推测N-酰基高丝氨酸内酯酶为亲水性蛋白。
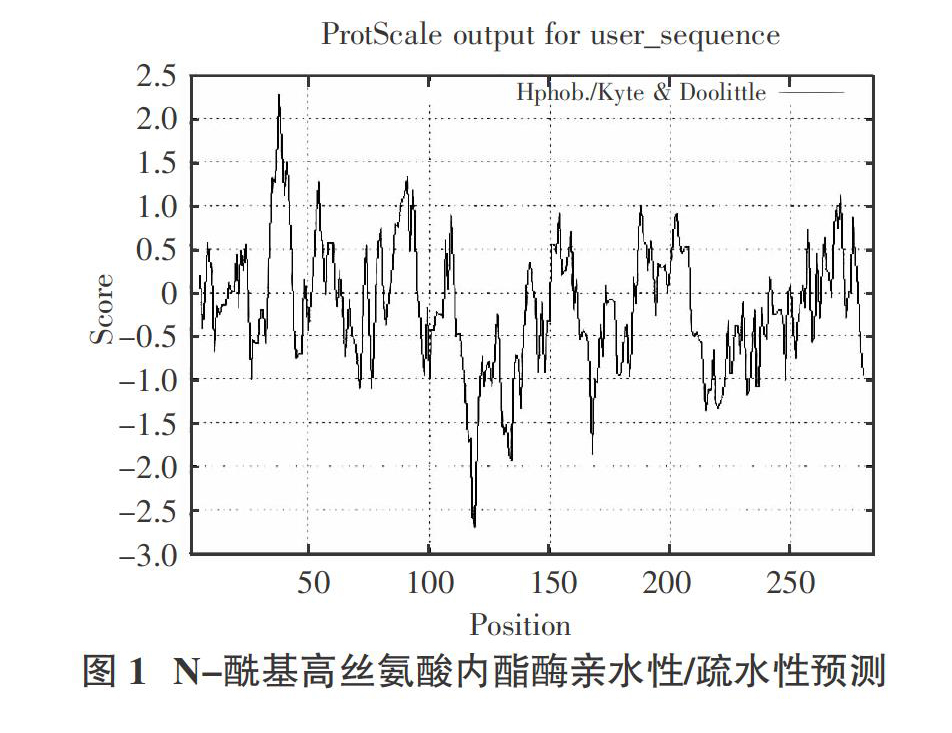
2.2? ?N-酰基高丝氨酸内酯酶亲水性/疏水性分析
利用ProtScale分析了N-酰基高丝氨酸内酯酶的亲水性/疏水性。通过图1可以看出,在N-酰基高丝氨酸内酯酶多肽链中,第119位谷氨酸有最低分值-2.689,亲水性最强;第38位异亮氨酸有最高分值2.267,疏水性最强。亲水性氨基酸数量较多,亲水性平均值为-0.187,推测N-酰基高丝氨酸内酯酶表现为亲水性。
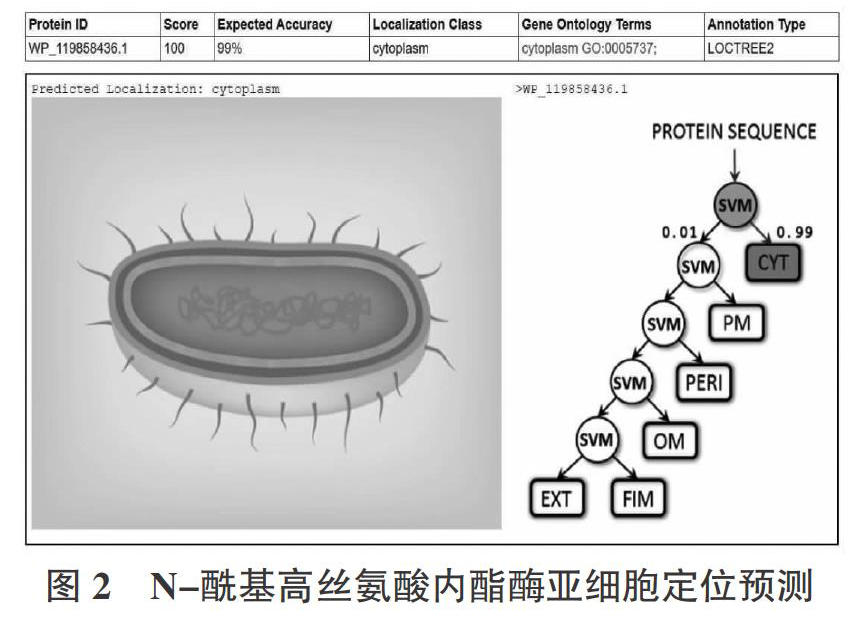
2.3? ?N-酰基高丝氨酸内酯酶亚细胞定位分析
PSORTb version3.0.2是目前最精确的细菌定位预测工具之一,利用PSORTb version 3.0.2分析了N-酰基高丝氨酸内酯酶的亚细胞定位。N-酰基高丝氨酸内酯酶的亚细胞定位分数分别为细胞质8.96,细胞质膜0.51,周质0.26,外膜0.01,细胞外0.26。定位分数显示N-酰基高丝氨酸内酯酶定位在细菌的细胞质中。利用Loc Tree3分析了N-酰基高丝氨酸内酯酶的亚细胞定位,结果如图2所示:N-酰基高丝氨酸内酯酶定位在细胞质中,亚细胞定位分数为100,精确度99%。基于上述2种在线工具的亚细胞定位预测表明,N-酰基高丝氨酸内酯酶定位在细胞质中。
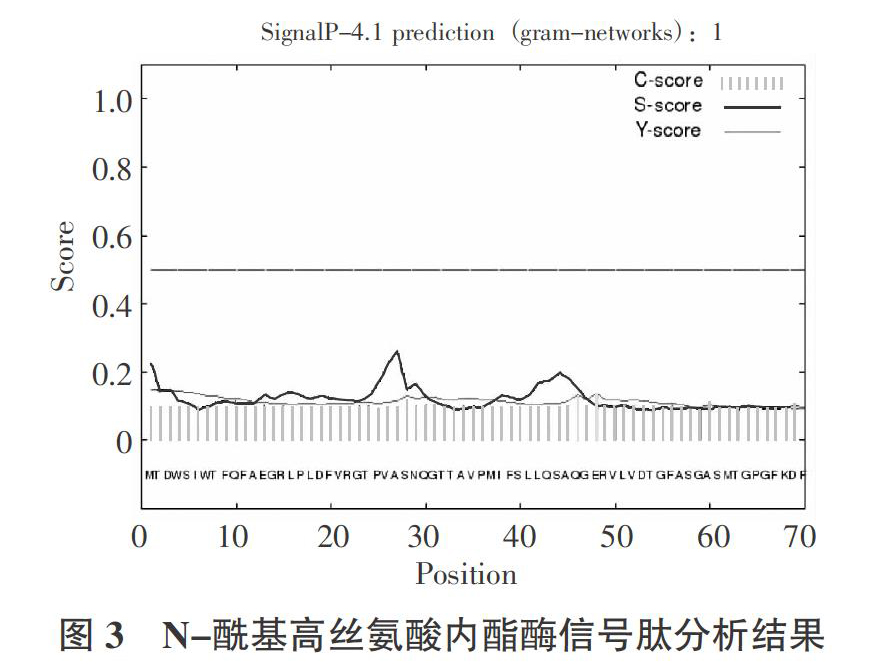
2.4? ?N-酰基高丝氨酸内酯酶信号肽分析
利用SignalP 4.1 Server分析了N-酰基高丝氨酸内酯酶的信号肽,结果如图3所示:原始剪切位点C最大值在第48个氨基酸,分值为0.132;综合剪切位点Y最大值在第48个氨基酸,分值是0.137;信号肽S最大值在第27个氨基酸,分值为0.262;平均信号肽S值为第1~47个氨基酸,平均值是0.134;D值为第1~47个氨基酸,平均值是0.136;临界值是0.570。D值是信号肽S最大值和综合剪切位点Y最大值的平均值,对区分是否为分泌蛋白具有至关重要的作用。D值小于临界值,未发现信号肽序列,故推断N-酰基高丝氨酸内酯酶为非分泌性蛋白。
2.5? ?N-酰基高丝氨酸内酯酶二级结构分析
利用SOPMA分析了N-酰基高丝氨酸内酯酶的二级结构,结果如图4所示:N-酰基高丝氨酸内酯酶可能包含的二级结构分别为α-螺旋(蓝色)67个,占23.59%;β-转角(绿色)28个,占9.86%;延伸链(红色)64个,占22.54%;无规则卷曲(紫色)125个,占44.01%。可见无规则卷曲是N-酰基高丝氨酸内酯酶二级结构中最大量的结构元件。
2.6? ?N-酰基高丝氨酸内酯酶三级结构分析
利用SWISS-MODEL分析了N-酰基高丝氨酸内酯酶三级结构,得到以PDB ID: 3aj3.1.A(4-吡哆醇内酯酶)为模板的预测模型,作为本研究的N-酰基高丝氨酸内酯酶预测模型。从图5看出,N-酰基高丝氨酸内酯酶预测模型与模板序列相似度为23.87%。N-酰基高丝氨酸内酯酶预测模型的全局模型质量评估分数(GMQE)为0.55,接近1。GMQE是一种质量评估方法,生成的GMQE在0到1之间,数字越高表明可靠性越高。通过SWISS-MODEL预测得出,N-酰基高丝氨酸内酯酶的三级结构(图6),可见其α-螺旋和无规则卷曲结构丰富,数量较多,推测α-螺旋和无规则卷曲对N-酰基高丝氨酸内酯酶生物学功能的发挥具有重要作用。
2.7? ?N-酰基高丝氨酸内酯酶磷酸化位点分析
利用NetPhosBac 1.0 server分析了N-酰基高丝氨酸内酯酶的磷酸化位点。从图7可以看出,磷酸化位点有6个,其中丝氨酸磷酸化位點包括73 S(残基得分0.621)、183 S(残基得分0.581)、194 S(残基得分0.525)、278 S(残基得分0.501),苏氨酸磷酸化位点包括24 T(残基得分0.510)、174 T(残基得分0.508)。残基得分介于0和1,当得分高于0.5时,残基被预测为磷酸化位点。
2.8? ?N-酰基高丝氨酸内酯酶跨膜结构域分析
利用TMHMM 2.0 server分析了N-酰基高丝氨酸内酯酶的跨膜结构域,结果如图8所示。预测N-酰基高丝氨酸内酯酶位于膜外的概率接近100%,位于膜内和跨膜区域的概率几乎为0。因N-酰基高丝氨酸内酯酶无跨膜结构域,所以图8中不显示相应标记。推测N-酰基高丝氨酸内酯酶不属于跨膜蛋白。
2.9? ?N-酰基高丝氨酸内酯酶功能结构域分析
利用NCBI的保守域数据库(CDD)进行功能结构域分析,N-酰基高丝氨酸内酯酶参与细菌群体感应淬灭。从图9可看出,N-酰基高丝氨酸内酯酶在31-251位氨基酸处含有保守结构域AHL_lactonase_MBL-fold,该结构域属于MBL折叠金属水解酶超家族,该家族主要由具有多种生物学功能的水解酶组成。N-酰基高丝氨酸内酯是许多革兰氏阴性细菌群体感应系统中负责协调基因表达的信号分子。N-酰基高丝氨酸内酯酶具有群体感应淬灭功能,催化N-酰基高丝氨酸内酯的高丝氨酸内酯环的水解和开环,从而阻断群体感应系统功能的发挥。
3? 小结与讨论
益生菌是指当以足够数量存在时可对机体健康产生有益作用的活的微生物[8 ]。沼泽红假单胞菌属于益生菌的一种,可作为微生物饲料添加剂使用,具有极高的应用价? ? ?值[9 ]。微生物饲料添加剂在高温、胃酸、胆盐等条件下易失活[10 ],生物被膜状态下的益生菌能够很好地解决上述诸多问题[11 - 12 ]。与常规益生菌制剂相比,在高密度生物被膜状态下益生菌制备的制剂具有更好的耐热性、耐酸性和抗冷冻干燥能力[13 ]。我们利用NCBI的保守域数据库进行功能结构域分析,结果显示N-酰基高丝氨酸内酯酶参与细菌群体感应淬灭。群体感应是微生物之间通过信号分子进行交流的一种通讯机制,可使细菌表现出不同的生理行为,例如生物被膜的形成与生长[14 - 16 ]。细菌群体感应淬灭具有3种途径,一是阻碍信号分子合成蛋白的产生,从源头上使信号分子无法生成;二是通过降解信号分子,使信号分子浓度处于阈值以下,阻止下游基因的表达;三是阻止信号分子与相应受体蛋白的结合,使其无法行使转录调节因子的功能[6,17 - 18 ]。群体感应淬灭酶是最主要的群体感应淬灭手段之一,通过降解信号分子以及阻断信号的产生和识别起到淬灭的作用。N-酰基高丝氨酸内酯都有一个高丝氨酸内酯环和酰胺键,可为群体感应淬灭酶提供相应的酶切位点。根据酶切位点的不同,群体感应淬灭酶包括内酯酶、酰胺酶和氧化还原酶三大类,其中N-酰基高丝氨酸内酯酶表现出了极强的淬灭活性。N-酰基高丝氨酸内酯酶是群体感应淬灭酶中首个被确定的群体感应淬灭酶[6,19 ]。N-酰基高丝氨酸内酯酶能够水解N-酰基高丝氨酸内酯的内酯环,产生N-酰基高丝氨酸。在酸性条件下,N-酰基高丝氨酸能够重新环化成N-酰基高丝氨酸内酯,因此N-酰基高丝氨酸内酯酶的淬灭作用是可逆的。N-酰基高丝氨酸内酯酶可分为不同的蛋白家族,不同的蛋白家族又包含多种具有同源性的酶[20 ],本研究中的N-酰基高丝氨酸内酯酶属于金属水解酶超家族,具有水解酶活性,能够催化N-酰基高丝氨酸内酯的高丝氨酸内酯环的水解和开环。N-酰基高丝氨酸内酯酶通过降解革兰氏阴性细菌群体感应系统信号分子,从而调控细菌生物被膜的形成。N-酰基高丝氨酸内酯酶作为生物被膜形成过程中的关键调控蛋白,是理想的生物被膜增强剂的候选蛋白,因此,通过不断的对其进行深入研究和分析,对生物被膜的人为控制将具有非常诱人的前景。
参考文献:
[1] 易洪斌,丁松林,张海波,等.? 日粮中添加沼泽红假单胞菌对生长牛生长性能和血液生理生化指标的影响[J].? 中国畜牧杂志,2020,
56(9):128-132.
[2] 李美洁,夏青青,HARWOOD C S,等.? 沼泽红假单胞菌作为微生物细胞工厂的应用[J].? 生物加工过程,2020,18(1):27-34.
[3] SALAS-JARA M J,ILABACA A,VEGA M, et al.? Biofilm forming lactobacillus:new challenges for the development of probiotics[J].? Microorganisms,2016,4(3):35.
[4] 崔天琦,白凤翎,励建荣.? 基于AHLs介导的革兰氏阴性菌群体感应调控及淬灭机制研究进展[J].? 中国食品学报,2020,20(8):308-320.
[5] 周? ?朋,王? ?喆,包美娇,等.? 微生物群体感应系统的调控机制及应用研究进展[J].? 生物加工过程,2019,17(3):236-243.
[6] 常? ?晶,史国萃,曾名湧,等.? 细菌群体感应淬灭酶及其应用研究进展[J].? 生物加工过程,2019,17(3):244-250.
[7] SIKDAR R,ELIAS M. Quorum quenching enzymes and their effects on virulence,biofilm,and microbiomes:a review of recent advances[J].? Expert Review of Anti-infective Therapy,2020,8(4):11-13.
[8] 刘? ?政,高爱琴,张铁鹰,等.? 益生菌制剂作为动物饲料添加剂的研究进展[J].? 饲料研究,2020,43(8):148-151.
[9] 田启文,郭? ?振,嵇乐乐,等.? 水产养殖中益生菌研究进展[J].? 工业微生物,2019,49(4):50-55.
[10] 修建成.? 微生物饲料添加剂的研究与应用[J].? 饲料博览,2015(11):20-23.
[11] 靳胜男,舒慧萍,张冬星,等.? 猪源生物被膜乳酸菌的筛选、鉴定与体外益生特性评价[J].? 河南农业科学,2019,48(12):121-127.
[12] 靳胜男,舒慧萍,张冬星,等.? 猪源生物被膜乳酸菌的筛选与体外益生特性评价[J].? 中国兽医科学,2019,49(10):1288-1296.
[13] 刘? ?蕾,刘? ?义,李平兰.? 益生菌生物被膜的研究进展[J].? 食品科学,2016,37(9):214-219.
[14] 张天震,刘伶普,李文超,等.? 群体感应系统介导细菌生物膜形成的研究进展[J].? 生物加工过程,2020,18(2):177-183.
[15] 肖梦圆,武瑞赟,谭春明,等.? 群体感应系统及其抑制剂对细菌生物被膜调控的研究进展[J].? 食品科学,2020,41(13):227-234.
[16] 张炼辉.? 微生物群体感应系统的研究进展[J].? 华南农业大学学报,2019,40(5):50-58.
[17] 王志航,冯? ?雪,李树仁,等.? 细菌群体感应通讯系统淬灭及应用[J].? 药物生物术,2018,
25(5):443-447.
[18] 王? ?岩,于雅萌,张静静,等.? 海洋微生物群体感应与群体感应淬灭的开发利用[J].? 生物资源, 2017,39(6):413-422.
[19] 孙? ?锋,严慧聪,汪美贞.? 细菌群体感应调控多样性及群体感应淬灭[J].? 微生物學报,2019,59(3):454-467.
[20] 范兴辉,王惠杉,何杰华,等.? 细菌群体感应淬灭酶及其病害防治研究进展[J].? 生物技术通报,2017,33(10):80-87.
(本文责编:陈? ? 伟)
