抗菌肽在母猪生产中的应用及作用机制
问鑫 高凤仙
摘要:抗菌肽作为抗生素替代品的首选,得到了大量的研究。本文详细的综述了抗菌肽杀菌的作用机理,分别阐述了抗菌肽的作用机制及对母猪生产性能的影响。
关键字:抗菌肽;作用机理;膜损伤;母猪;生产性能
抗菌肽(antimicrobial peptides,AMPs)是指在外界条件作用的刺激下由特定的基因编码、由核糖体合成的一类可抵御外界微生物侵害、清除体内抗微生物的小分子阳离子多肽,是动物非特异性免疫系统的重要效应分子。本文综述了抗菌肽的结构特点、理化特性、作用机制及其在母猪生产中的应用。
1抗菌肽的结构特点与理化性质
目前,已分离或预测出的抗菌肽超过1200种,但大部分抗菌肽只包含50或更少的氨基酸残基,最短的抗菌肽仅有6个残基,分子量较小。其N端含有赖氨酸、精氨酸等亲水性氨基酸残基,C端则富含丙氨酸、甘氨酸、缬氨酸等疏水性氨基酸残基,这使得抗菌肽有水脂两亲性结构;同时,可赖氨酸、精氨酸等碱性氨基酸通常会使抗菌肽等电点大于7,表现出一定的阳离子特征,可与两性分子或电负性细胞膜结合,与细菌的细胞膜发生作用。抗菌肽的中间部分含有大量的脯氨酸和二硫键,由此易形成α-螺旋、β-折叠、伸展片层结构和环形等与其抗菌活性密切相关的结构。
作为抗生素替代物的首选。抗菌肽具有以下特点:(1)广谱抗菌活性:抗菌肽不仅能杀灭革兰氏阳性、阴性菌、真菌、病毒、原虫,并对肿瘤和癌细胞有一定的杀伤作用:(2)热稳定性:抗菌肽具有较强的耐热性,在沸水中30min仍可保持较高的抗菌活性,可耐受饲料制粒的高温过程:(3)高效性:抗菌肽对病原微生物的最小抑菌浓度(MIC)仅为0.1~10μg/ml,并且可在极短的时间内杀灭所有病原体:(4)特异性:尽管抗菌肽是动物先天性防御系统的产物,但是具有特异性,只对原核生物细胞和病变的真核细胞产生作用,对其他微生物和健康的真核细胞无杀伤作用;(5)抗水解:有些抗菌肽可抵抗胰蛋白酶或胃蛋白酶的水解,使其可在动物体内完整的发挥作用。此外,抗菌肽还有水溶性好、不残留、不易产生耐药性等特点。
2抗菌肽对母猪的影响及其作用机理
随着近年来养猪生产的集约化、规模化,养殖户将大量精力放在了提高生长和产肉性能等方面上,使母猪长期处于应激状态中,引起母猪的内分泌机能失调、食欲不振、体脂含量降低等应激反应,并加剧了母猪泌乳障碍,直接影响了仔猪的成活率和生长性能,造成了巨大的经济损失。作为抗生素替代品的首选,将抗菌肽加入母猪日粮中,可提高母猪的生产性能。
2.1抑杀影响母猪生产的病毒
猪呼吸与繁殖综合症(PRRSV)和猪圆环病毒病(PCV-2)是影响我国养猪业发展的两个巨大障碍,其可使猪群生产性能下降,母猪流产增多。万小平等(2011)采用融合抗菌肽MAPWA和PGBD-2加入后备母猪日粮中,研究发现,融合抗菌肽MAPWA和PGBD-2可有效减少体内PCV-2、PRRSV的带毒率;处理后备母猪血液PCV-2、PRRSV病毒含量也明显减少。显示饲喂融合抗菌肽MAPWA和PGBD-2产生了良好的抑制及清除病毒的作用,对降低后备母猪体内PCV-2、PRRSV带毒率有明显成效。说明抗菌肽具有防治常见疾病的前景。此外,有研究表明,抗菌肽对提高猪瘟抗体水平,改善抗体均匀度方面具有十分显著的效果,充分验证了抗菌肽对调节动物免疫机能、增强动物免疫力方面的作用。
其增强母猪免疫力的主要机理为:抗菌肽可抑制病毒DNA合成的过程,还可通过阻碍其复制、转录,影响蛋白质的翻译过程,间接的抑制病毒蛋白质合成。王军和庞广昌(2005)报道,抗菌肽Attacin可干扰大肠杆菌的膜蛋白转录,阻碍蛋白质合成,从而增加细胞膜的通透性。杀灭细菌。抗菌肽Indolicidin可通过与脂多糖的相互结合,达到抑制蛋白质合成的效果。ZHOU和CHEN(2011)研究了抗菌肽apidaecins的亚型apidaecin IB与大肠杆菌膜蛋白的作用。发现大肠杆菌细胞培养过程中加入apidaecin IB,可使维持膜脂平衡的ftsH过量分泌。阻碍了膜蛋白合成的第一个关键步骤,抑制其合成。Yenugu等(2004)报道,抗菌肽HE2的几个亚型(HE2al-pha、HE2betal和HE2beta2)不但可与大肠杆菌细胞膜作用使胞内物质泄露,还可抑制其DNA、RNA和蛋白质的合成。
刘立伟和邓磊(2012)报道,抗菌肽能刺激细胞,使细胞内的溶酶体和内质网中的物质如钙离子和蛋白酶排入细胞质中,进一步刺激线粒体释放细胞色素C,通过一系列酶促反应诱导细胞凋亡。有些抗菌素如thanatin、histatin还可抑制细胞呼吸作用。進而抑杀细菌。此外,LIU等(2013)采用包含诱导表达的抗菌肽LL-37的穿梭载体,研究大肠杆菌要有氧或无氧条件下的生长状况,研究发现。LL-37会使大肠杆菌生长受阻,使细胞形态变为丝状;同时,LL-37不仅可提高其在细胞内的活性氧含量,在有氧条件下引起致死膜去极化,增加胞膜的通透性,并且通过转录分析发现LL-37可通过阻碍氧化磷酸化相关基因的表达,影响细胞糖代谢和能量转化的过程以及细胞内离子的氧化还原状态。由此提出了一种新的靶向能量代谢的AMPs受体介导的杀菌机制。
2.2降低母猪死产率,提高仔猪成活率
皮灿辉等(2008a)用抗菌肽替代猪场使用的抗生素饲喂母猪和其所产仔猪,并对508头母猪和它们分娩产生的5245头猪仔进行追踪统计并记录其健康状况。结果表明,试验组木乃伊死胎率仅为0.31%,比对照组(7.15%)降低了22倍。在哺乳阶段,未使用抗菌肽饲料添加剂的对照组死亡率(2.36%)是试验组(0.35%)的7倍。在保育阶段,未使用抗菌肽饲料添加剂的对照组死亡率(3.76%)是试验组(1.32%)的3倍。潘行正等(2010)研究了抗菌肽对减少母猪死胎率的效果,结果表明,添加抗菌肽制剂的母猪试验组死产率为5.77%,按日常用药方案使用抗生素的对照组死产率为12.75%:其中试验组木乃伊胎的死产率仅为0.31%。对照组却达到7.16%。添加抗菌肽制剂的试验组仔猪在哺乳期和保育期的死亡率分别为0.34%、1.31%,同期对照组分别为2.35%、3.65%;对照组死亡率显著高于试验组。可见抗菌肽制剂能显著降低母猪的死产率与提高仔猪的成活率与猪只的生产性能和健康水平。
抗菌肽之所以能降低母猪死产率与提高仔猪成活率,与其广谱的抗菌作用有关:细菌细胞壁的主要成分是肽聚糖,是维持细胞强度和形状的外被,具有保护细胞形态的作用。当细胞壁被破坏,细菌的形态及对渗透压的敏感性都会产生变化,引起细胞死亡。抗菌肽可通过破坏细胞壁和抑制细胞壁合成两方面抑杀细菌。
Herusimenka等(2005)研究抗菌肽与细菌的相互作用时指出,二者之间的相互作用主要通过肽与多糖之间实现的。抗菌肽带正电荷,可取代细胞壁表面的镁离子,与细菌细胞壁上的负电荷物质通过静电引力结合,引起细胞壁结构发生变化,形成细胞壁紊乱区域,促使抗菌肽穿过细胞壁。接近细胞膜。在革兰氏阴性菌中,带负电荷的物质为脂多糖和荚膜多糖,而在革兰氏阳性菌的细胞壁表面有带负电荷的是磷壁酸。唐亚丽(2009)用抗菌肽MDpep5和MDpep9处理了大肠杆菌、金黄色葡萄球菌,结果表明,两种细菌的A630nm值均有不同程度的下降,说明家蝇抗菌肽对细菌细胞壁有一定的溶解作用,可使细菌细胞壁的通透性增加。乔媛媛等(2010)利用电子显微镜观察了抗菌肽的作用机理,研究表明。抗菌肽作用于大肠杆菌12h后,细菌的质壁发生分离。使细胞壁出现空泡。进一步证明了抗菌肽可破坏细胞壁。宫霞等(2011)指出,抗菌肽MDL-2可与细菌细胞壁,使其溶解,对革兰氏阳性细菌金黄色葡萄球菌的作用在较低浓度下即可发生,但对革兰氏阴性菌的作用则需达到一定的浓度才可破坏其细胞壁。这可能与两种菌表面的不同组分有关,在抗菌肽达到一定浓度时才能溶解革兰氏阴性菌表面的脂多糖。
抗菌肽不仅可破坏、溶解细胞壁,并通过抑制细胞壁的合成来表达其抗菌活性。蔡永峰和彭德轩(2000)报道,杆菌肽锌可抑制细菌细胞壁合成的脱磷酸化过程,阻碍磷脂载体的运转及向细胞壁支架输送粘肽,从而抑制细胞壁的合成。Haster等(2006)研究发现,Lipid Ⅱ是细胞壁的合成前体。而抗菌肽Nisin可移除Lipid Ⅱ在细胞壁上的合成位点。从而阻碍细胞壁的合成。Harder(2001)等指出。抗菌肽β-defensin3可抑制细菌细胞壁的形成,使细菌的细胞形态异常、阻碍细菌分裂,导致细胞壁穿孔。引起细菌死亡。说明抗菌肽可通过阻断细胞壁的合成过程或移除其合成所需的物质来达到抑制细菌细胞壁合成的作用。
2.3提高妊娠母猪与仔猪的免疫力,节约生产成本
王阿荣(2011)将天蚕素抗菌肽加入母猪日粮中,研究发现。添加天蚕素抗菌肽对母猪的繁殖力和仔猪的生长性能等均无显著影响(P>0.05),但是试验Ⅲ组在这两项指标上较对照组都有提高的趋势:各试验组母猪初乳中的水分含量均高于对照组,但差异不显著(P>0.05)。各试验组母猪初乳中乳糖含量均高于对照组,其中只有试验Ⅱ组与对照组差异显著(P <0.05),其它试验组较对照组差异不显著(P>0.05)。各试验组母猪初乳中氨基酸水平和脂肪酸含量与对照组相比均无显著差异(P>0.05)。从试验整体结果看。在母猪日粮中添加60g/t天蚕素抗菌肽的试验Ⅲ组效果最佳。说明抗菌肽饲料添加剂可有效提高母猪的生产性能。并有效地降低饲养成本,为养殖户带来巨大的经济效益。
由于抗菌肽提高了母猪和仔猪的免疫力,在生產中的用药及人力上有了巨大的节约,可以进一步为养殖户带来可观的效益。在这里,笔者将为大家普及抗菌肽的几种抗菌模式。使养殖户更加明了的认知抗菌肽。
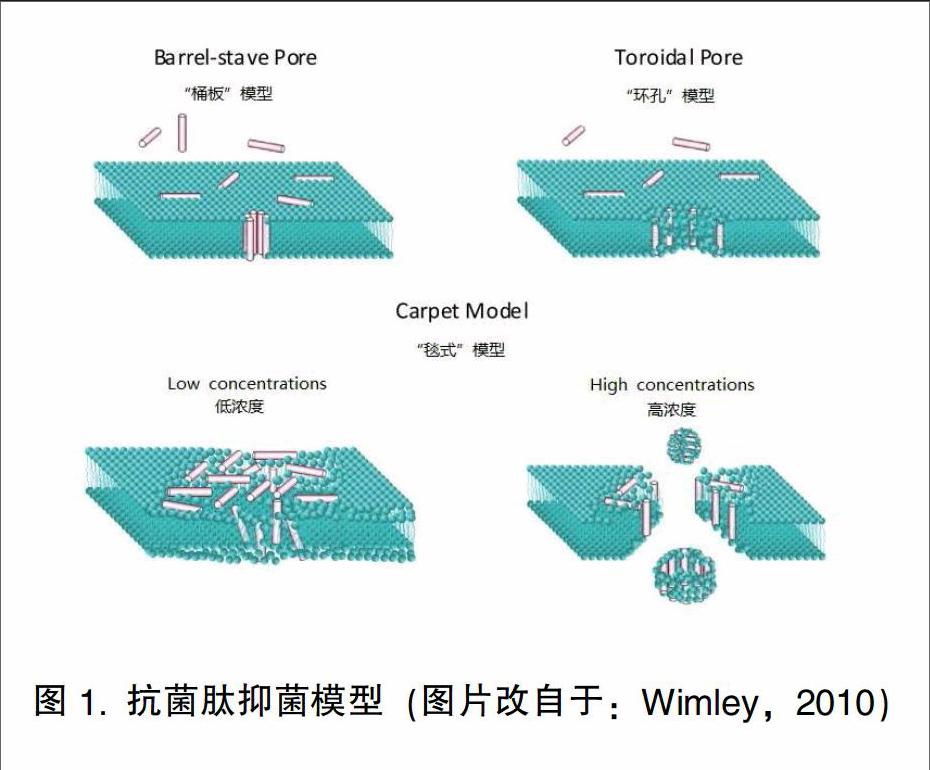
2.3.1“桶板”模型(Barrel-stave model)
在该模型中,抗菌肽通过静电引力聚集在胞膜表面,形成肽聚体,然后肽聚体的疏水部分垂直插入磷脂双分子层破坏细胞膜,形成离子通道,使细胞渗透压变化、胞质泄露,导致细胞死亡。唐亚丽(2009)指出,通过该模型产生抗菌作用的抗菌肽即可以单体形式,也可以多体形式与胞膜结合:当单体在膜表面的密度较低时,可互相识别、聚合,形成多聚体;多肽和胞膜的疏水部分相互结合。形成离子通道:在离子通道形成过程中,可通过在“桶板”上增加单体及小多体,扩大通道,加速细胞死亡。Gkeka和Sark-isov(2009)报道,抗菌肽跨膜形成的“桶板”通道的主要作用为α-螺旋。每6个α-螺旋结构的肽可形成内径0.52nm左右的螺旋肽,并由此形成“桶板”状的通道,其中孔道内侧为肽的亲水部分,外侧为疏水部分。该模型说明抗菌肽具有两亲性,并说明具有两亲性的α-螺旋结构的抗菌肽是形成膜孔洞的关键因素。另外,刘忠渊(2008a)指出,除了具有双亲性的α-螺旋,疏水性的α-螺旋或B折叠结构也是使抗菌肽具有穿膜孔洞作用的必需结构。
2.3.2“环孔”模型(Toroidal Dore model)
“环孔”模型最初是由Ludtke等在研究滑爪蟾素的作用机理时提出的,滑爪蟾素具有两性α-螺旋结构。与胞膜作用,可形成透膜孔道(刘立伟和邓磊,2012)。与“桶板”模型不同的是,抗菌肽与膜结合时形成明显的α-螺旋结构,肽的亲水部分与脂质的亲水部分结合,共同形成跨膜通道。当肽与磷脂的比例达到一定的值,抗菌肽互相结合。使膜表面的曲面张力变化,导致脂质双分子层内外两层贯通,改变胞膜结构,从而形成环孔状通道,最终引起细胞死亡。Bozelli等(2012)通过圆二色和荧光光谱对含13个残基的阳离子抗菌肽TRP3和含不同脂质的胞膜模型之间的作用进行了研究,用大单层脂质体(LUV)模拟肽一脂质聚合体、用胶束模仿“环孔”模型的拓扑结构,结果表明,只有当双分子层有负电荷的磷脂,TRP3才能与其结合,而无论胶束表面电荷的正负。TRP3均可与之结合,说明静电作用在“环孔”模型中不占主导作用,并指出。脂质双分子层的曲率可影响肽的二级结构。Sobko等(2004)研究了大肠杆菌素E1(col-icinEl)在磷脂双分子层和脂质体上形成环孔通道的活性,指出colieinEl形成环孔通道的活性取决于脂质双分子层的曲率,说明脂质直接参与了eolieinEl诱导孔壁形成的过程。并报道col-icinEl通过形成环孔通道。导致细胞凋亡。Yoneyama等(2009)报道,lacticin Q是一类可在细胞膜上形成孔隙的细菌素,并且是目前第一个可使蛋白泄露的抗菌肽。其形成孔隙的过程如下:首先,快速与细胞外膜结合:其次,形成至少4.6nm的孔隙,引起蛋白质泄露:再次,lac-ticin Q从细胞膜外侧迁移至膜内侧。
2.3.3“毯式”模型(Carpet model)
Pouny等(1992)在研究抗菌肽dermaseptin与磷脂膜的相互作用时提出了抗菌肽的“毯式”模型。当浓度较低时阳离子抗菌肽分子以“平铺”的方式覆盖在带负电的细胞膜表面,平行于磷脂双分子层。使其疏水部分与细胞膜结合:当抗菌肽的浓度达到一定的阈值,抗菌肽互相结合形成肽聚体,肽聚体可与细胞膜结合,形成类似洗涤剂一脂质混合物,使完整的细胞膜迅速崩溃、细胞膜被溶解,造成细胞内容物流失、细胞死亡(Wimley,2010;刘忠渊,2008a)。Lopez-Oyama等(2011)对抗菌肽Bactenecin与细胞膜的相互作用进行探究的过程中发现,使抗菌肽通过“毯式”模型产生抗菌作用的结构主要是β-折叠结构,这可能是由于β-折叠所特有的长丝状网格是与细胞膜产生作用的主要结构,Fernandez等(2012)分别用DMPC(二肉豆蔻酰磷脂酰胆碱)和DMPC/DMPG(二肉豆蔻酰磷脂酰甘油)作为真核和原核的模型膜,研究了抗菌肽aurein1.2与胞膜的相互作用,结果表明,anrein1.2与两种膜表面均有较强的相互作用,进而导致脂质双分子层的破坏和膜溶解,并根据不同类型的磷脂膜的结构变化,说明该抗菌肽与膜作用的机制为“毯式”模型(如图1所示)。
4展望
抗菌肽作为一种功能性添加剂用于分娩前后的母猪饲粮时,可以有效提高母猪的免疫机能和死产率、仔猪初生重和断奶重,显著降低哺乳期母猪产生的各种应激以及仔猪的腹泻率和死亡率,进而提高整体养殖效益。对于1000头母猪场而言,使用抗菌肽饲料添加剂可使年经济效益至少增加59.25万元。
但有关于抗菌肽的研究还有以下问题有待解决:
(1)抗菌肽是动植物免疫系统受到刺激后产生的物质。含量极微:虽然可利用化学方法合成,但成本较高,无法进行大规模的生产;(2)抗菌肽种类很多,有广谱的抗菌性:要发现一种生产成本低,使用效果好的抗菌肽添加剂,还需不断的研究探索;(3)抗菌肽种类丰富,抗菌机理各不相同,目前仍没有非常明确的、统一的理论;这急需研究者不断总结归纳抗菌肽的作用机制,加快抗菌肽应用的推广。
摘要:抗菌肽作为抗生素替代品的首选,得到了大量的研究。本文详细的综述了抗菌肽杀菌的作用机理,分别阐述了抗菌肽的作用机制及对母猪生产性能的影响。
关键字:抗菌肽;作用机理;膜损伤;母猪;生产性能
抗菌肽(antimicrobial peptides,AMPs)是指在外界条件作用的刺激下由特定的基因编码、由核糖体合成的一类可抵御外界微生物侵害、清除体内抗微生物的小分子阳离子多肽,是动物非特异性免疫系统的重要效应分子。本文综述了抗菌肽的结构特点、理化特性、作用机制及其在母猪生产中的应用。
1抗菌肽的结构特点与理化性质
目前,已分离或预测出的抗菌肽超过1200种,但大部分抗菌肽只包含50或更少的氨基酸残基,最短的抗菌肽仅有6个残基,分子量较小。其N端含有赖氨酸、精氨酸等亲水性氨基酸残基,C端则富含丙氨酸、甘氨酸、缬氨酸等疏水性氨基酸残基,这使得抗菌肽有水脂两亲性结构;同时,可赖氨酸、精氨酸等碱性氨基酸通常会使抗菌肽等电点大于7,表现出一定的阳离子特征,可与两性分子或电负性细胞膜结合,与细菌的细胞膜发生作用。抗菌肽的中间部分含有大量的脯氨酸和二硫键,由此易形成α-螺旋、β-折叠、伸展片层结构和环形等与其抗菌活性密切相关的结构。
作为抗生素替代物的首选。抗菌肽具有以下特点:(1)广谱抗菌活性:抗菌肽不仅能杀灭革兰氏阳性、阴性菌、真菌、病毒、原虫,并对肿瘤和癌细胞有一定的杀伤作用:(2)热稳定性:抗菌肽具有较强的耐热性,在沸水中30min仍可保持较高的抗菌活性,可耐受饲料制粒的高温过程:(3)高效性:抗菌肽对病原微生物的最小抑菌浓度(MIC)仅为0.1~10μg/ml,并且可在极短的时间内杀灭所有病原体:(4)特异性:尽管抗菌肽是动物先天性防御系统的产物,但是具有特异性,只对原核生物细胞和病变的真核细胞产生作用,对其他微生物和健康的真核细胞无杀伤作用;(5)抗水解:有些抗菌肽可抵抗胰蛋白酶或胃蛋白酶的水解,使其可在动物体内完整的发挥作用。此外,抗菌肽还有水溶性好、不残留、不易产生耐药性等特点。
2抗菌肽对母猪的影响及其作用机理
随着近年来养猪生产的集约化、规模化,养殖户将大量精力放在了提高生长和产肉性能等方面上,使母猪长期处于应激状态中,引起母猪的内分泌机能失调、食欲不振、体脂含量降低等应激反应,并加剧了母猪泌乳障碍,直接影响了仔猪的成活率和生长性能,造成了巨大的经济损失。作为抗生素替代品的首选,将抗菌肽加入母猪日粮中,可提高母猪的生产性能。
2.1抑杀影响母猪生产的病毒
猪呼吸与繁殖综合症(PRRSV)和猪圆环病毒病(PCV-2)是影响我国养猪业发展的两个巨大障碍,其可使猪群生产性能下降,母猪流产增多。万小平等(2011)采用融合抗菌肽MAPWA和PGBD-2加入后备母猪日粮中,研究发现,融合抗菌肽MAPWA和PGBD-2可有效减少体内PCV-2、PRRSV的带毒率;处理后备母猪血液PCV-2、PRRSV病毒含量也明显减少。显示饲喂融合抗菌肽MAPWA和PGBD-2产生了良好的抑制及清除病毒的作用,对降低后备母猪体内PCV-2、PRRSV带毒率有明显成效。说明抗菌肽具有防治常见疾病的前景。此外,有研究表明,抗菌肽对提高猪瘟抗体水平,改善抗体均匀度方面具有十分显著的效果,充分验证了抗菌肽对调节动物免疫机能、增强动物免疫力方面的作用。
其增强母猪免疫力的主要机理为:抗菌肽可抑制病毒DNA合成的过程,还可通过阻碍其复制、转录,影响蛋白质的翻译过程,间接的抑制病毒蛋白质合成。王军和庞广昌(2005)报道,抗菌肽Attacin可干扰大肠杆菌的膜蛋白转录,阻碍蛋白质合成,从而增加细胞膜的通透性。杀灭细菌。抗菌肽Indolicidin可通过与脂多糖的相互结合,达到抑制蛋白质合成的效果。ZHOU和CHEN(2011)研究了抗菌肽apidaecins的亚型apidaecin IB与大肠杆菌膜蛋白的作用。发现大肠杆菌细胞培养过程中加入apidaecin IB,可使维持膜脂平衡的ftsH过量分泌。阻碍了膜蛋白合成的第一个关键步骤,抑制其合成。Yenugu等(2004)报道,抗菌肽HE2的几个亚型(HE2al-pha、HE2betal和HE2beta2)不但可与大肠杆菌细胞膜作用使胞内物质泄露,还可抑制其DNA、RNA和蛋白质的合成。
刘立伟和邓磊(2012)报道,抗菌肽能刺激细胞,使细胞内的溶酶体和内质网中的物质如钙离子和蛋白酶排入细胞质中,进一步刺激线粒体释放细胞色素C,通过一系列酶促反应诱导细胞凋亡。有些抗菌素如thanatin、histatin还可抑制细胞呼吸作用。進而抑杀细菌。此外,LIU等(2013)采用包含诱导表达的抗菌肽LL-37的穿梭载体,研究大肠杆菌要有氧或无氧条件下的生长状况,研究发现。LL-37会使大肠杆菌生长受阻,使细胞形态变为丝状;同时,LL-37不仅可提高其在细胞内的活性氧含量,在有氧条件下引起致死膜去极化,增加胞膜的通透性,并且通过转录分析发现LL-37可通过阻碍氧化磷酸化相关基因的表达,影响细胞糖代谢和能量转化的过程以及细胞内离子的氧化还原状态。由此提出了一种新的靶向能量代谢的AMPs受体介导的杀菌机制。
2.2降低母猪死产率,提高仔猪成活率
皮灿辉等(2008a)用抗菌肽替代猪场使用的抗生素饲喂母猪和其所产仔猪,并对508头母猪和它们分娩产生的5245头猪仔进行追踪统计并记录其健康状况。结果表明,试验组木乃伊死胎率仅为0.31%,比对照组(7.15%)降低了22倍。在哺乳阶段,未使用抗菌肽饲料添加剂的对照组死亡率(2.36%)是试验组(0.35%)的7倍。在保育阶段,未使用抗菌肽饲料添加剂的对照组死亡率(3.76%)是试验组(1.32%)的3倍。潘行正等(2010)研究了抗菌肽对减少母猪死胎率的效果,结果表明,添加抗菌肽制剂的母猪试验组死产率为5.77%,按日常用药方案使用抗生素的对照组死产率为12.75%:其中试验组木乃伊胎的死产率仅为0.31%。对照组却达到7.16%。添加抗菌肽制剂的试验组仔猪在哺乳期和保育期的死亡率分别为0.34%、1.31%,同期对照组分别为2.35%、3.65%;对照组死亡率显著高于试验组。可见抗菌肽制剂能显著降低母猪的死产率与提高仔猪的成活率与猪只的生产性能和健康水平。
抗菌肽之所以能降低母猪死产率与提高仔猪成活率,与其广谱的抗菌作用有关:细菌细胞壁的主要成分是肽聚糖,是维持细胞强度和形状的外被,具有保护细胞形态的作用。当细胞壁被破坏,细菌的形态及对渗透压的敏感性都会产生变化,引起细胞死亡。抗菌肽可通过破坏细胞壁和抑制细胞壁合成两方面抑杀细菌。
Herusimenka等(2005)研究抗菌肽与细菌的相互作用时指出,二者之间的相互作用主要通过肽与多糖之间实现的。抗菌肽带正电荷,可取代细胞壁表面的镁离子,与细菌细胞壁上的负电荷物质通过静电引力结合,引起细胞壁结构发生变化,形成细胞壁紊乱区域,促使抗菌肽穿过细胞壁。接近细胞膜。在革兰氏阴性菌中,带负电荷的物质为脂多糖和荚膜多糖,而在革兰氏阳性菌的细胞壁表面有带负电荷的是磷壁酸。唐亚丽(2009)用抗菌肽MDpep5和MDpep9处理了大肠杆菌、金黄色葡萄球菌,结果表明,两种细菌的A630nm值均有不同程度的下降,说明家蝇抗菌肽对细菌细胞壁有一定的溶解作用,可使细菌细胞壁的通透性增加。乔媛媛等(2010)利用电子显微镜观察了抗菌肽的作用机理,研究表明。抗菌肽作用于大肠杆菌12h后,细菌的质壁发生分离。使细胞壁出现空泡。进一步证明了抗菌肽可破坏细胞壁。宫霞等(2011)指出,抗菌肽MDL-2可与细菌细胞壁,使其溶解,对革兰氏阳性细菌金黄色葡萄球菌的作用在较低浓度下即可发生,但对革兰氏阴性菌的作用则需达到一定的浓度才可破坏其细胞壁。这可能与两种菌表面的不同组分有关,在抗菌肽达到一定浓度时才能溶解革兰氏阴性菌表面的脂多糖。
抗菌肽不仅可破坏、溶解细胞壁,并通过抑制细胞壁的合成来表达其抗菌活性。蔡永峰和彭德轩(2000)报道,杆菌肽锌可抑制细菌细胞壁合成的脱磷酸化过程,阻碍磷脂载体的运转及向细胞壁支架输送粘肽,从而抑制细胞壁的合成。Haster等(2006)研究发现,Lipid Ⅱ是细胞壁的合成前体。而抗菌肽Nisin可移除Lipid Ⅱ在细胞壁上的合成位点。从而阻碍细胞壁的合成。Harder(2001)等指出。抗菌肽β-defensin3可抑制细菌细胞壁的形成,使细菌的细胞形态异常、阻碍细菌分裂,导致细胞壁穿孔。引起细菌死亡。说明抗菌肽可通过阻断细胞壁的合成过程或移除其合成所需的物质来达到抑制细菌细胞壁合成的作用。
2.3提高妊娠母猪与仔猪的免疫力,节约生产成本
王阿荣(2011)将天蚕素抗菌肽加入母猪日粮中,研究发现。添加天蚕素抗菌肽对母猪的繁殖力和仔猪的生长性能等均无显著影响(P>0.05),但是试验Ⅲ组在这两项指标上较对照组都有提高的趋势:各试验组母猪初乳中的水分含量均高于对照组,但差异不显著(P>0.05)。各试验组母猪初乳中乳糖含量均高于对照组,其中只有试验Ⅱ组与对照组差异显著(P <0.05),其它试验组较对照组差异不显著(P>0.05)。各试验组母猪初乳中氨基酸水平和脂肪酸含量与对照组相比均无显著差异(P>0.05)。从试验整体结果看。在母猪日粮中添加60g/t天蚕素抗菌肽的试验Ⅲ组效果最佳。说明抗菌肽饲料添加剂可有效提高母猪的生产性能。并有效地降低饲养成本,为养殖户带来巨大的经济效益。
由于抗菌肽提高了母猪和仔猪的免疫力,在生產中的用药及人力上有了巨大的节约,可以进一步为养殖户带来可观的效益。在这里,笔者将为大家普及抗菌肽的几种抗菌模式。使养殖户更加明了的认知抗菌肽。
2.3.1“桶板”模型(Barrel-stave model)
在该模型中,抗菌肽通过静电引力聚集在胞膜表面,形成肽聚体,然后肽聚体的疏水部分垂直插入磷脂双分子层破坏细胞膜,形成离子通道,使细胞渗透压变化、胞质泄露,导致细胞死亡。唐亚丽(2009)指出,通过该模型产生抗菌作用的抗菌肽即可以单体形式,也可以多体形式与胞膜结合:当单体在膜表面的密度较低时,可互相识别、聚合,形成多聚体;多肽和胞膜的疏水部分相互结合。形成离子通道:在离子通道形成过程中,可通过在“桶板”上增加单体及小多体,扩大通道,加速细胞死亡。Gkeka和Sark-isov(2009)报道,抗菌肽跨膜形成的“桶板”通道的主要作用为α-螺旋。每6个α-螺旋结构的肽可形成内径0.52nm左右的螺旋肽,并由此形成“桶板”状的通道,其中孔道内侧为肽的亲水部分,外侧为疏水部分。该模型说明抗菌肽具有两亲性,并说明具有两亲性的α-螺旋结构的抗菌肽是形成膜孔洞的关键因素。另外,刘忠渊(2008a)指出,除了具有双亲性的α-螺旋,疏水性的α-螺旋或B折叠结构也是使抗菌肽具有穿膜孔洞作用的必需结构。
2.3.2“环孔”模型(Toroidal Dore model)
“环孔”模型最初是由Ludtke等在研究滑爪蟾素的作用机理时提出的,滑爪蟾素具有两性α-螺旋结构。与胞膜作用,可形成透膜孔道(刘立伟和邓磊,2012)。与“桶板”模型不同的是,抗菌肽与膜结合时形成明显的α-螺旋结构,肽的亲水部分与脂质的亲水部分结合,共同形成跨膜通道。当肽与磷脂的比例达到一定的值,抗菌肽互相结合。使膜表面的曲面张力变化,导致脂质双分子层内外两层贯通,改变胞膜结构,从而形成环孔状通道,最终引起细胞死亡。Bozelli等(2012)通过圆二色和荧光光谱对含13个残基的阳离子抗菌肽TRP3和含不同脂质的胞膜模型之间的作用进行了研究,用大单层脂质体(LUV)模拟肽一脂质聚合体、用胶束模仿“环孔”模型的拓扑结构,结果表明,只有当双分子层有负电荷的磷脂,TRP3才能与其结合,而无论胶束表面电荷的正负。TRP3均可与之结合,说明静电作用在“环孔”模型中不占主导作用,并指出。脂质双分子层的曲率可影响肽的二级结构。Sobko等(2004)研究了大肠杆菌素E1(col-icinEl)在磷脂双分子层和脂质体上形成环孔通道的活性,指出colieinEl形成环孔通道的活性取决于脂质双分子层的曲率,说明脂质直接参与了eolieinEl诱导孔壁形成的过程。并报道col-icinEl通过形成环孔通道。导致细胞凋亡。Yoneyama等(2009)报道,lacticin Q是一类可在细胞膜上形成孔隙的细菌素,并且是目前第一个可使蛋白泄露的抗菌肽。其形成孔隙的过程如下:首先,快速与细胞外膜结合:其次,形成至少4.6nm的孔隙,引起蛋白质泄露:再次,lac-ticin Q从细胞膜外侧迁移至膜内侧。
2.3.3“毯式”模型(Carpet model)
Pouny等(1992)在研究抗菌肽dermaseptin与磷脂膜的相互作用时提出了抗菌肽的“毯式”模型。当浓度较低时阳离子抗菌肽分子以“平铺”的方式覆盖在带负电的细胞膜表面,平行于磷脂双分子层。使其疏水部分与细胞膜结合:当抗菌肽的浓度达到一定的阈值,抗菌肽互相结合形成肽聚体,肽聚体可与细胞膜结合,形成类似洗涤剂一脂质混合物,使完整的细胞膜迅速崩溃、细胞膜被溶解,造成细胞内容物流失、细胞死亡(Wimley,2010;刘忠渊,2008a)。Lopez-Oyama等(2011)对抗菌肽Bactenecin与细胞膜的相互作用进行探究的过程中发现,使抗菌肽通过“毯式”模型产生抗菌作用的结构主要是β-折叠结构,这可能是由于β-折叠所特有的长丝状网格是与细胞膜产生作用的主要结构,Fernandez等(2012)分别用DMPC(二肉豆蔻酰磷脂酰胆碱)和DMPC/DMPG(二肉豆蔻酰磷脂酰甘油)作为真核和原核的模型膜,研究了抗菌肽aurein1.2与胞膜的相互作用,结果表明,anrein1.2与两种膜表面均有较强的相互作用,进而导致脂质双分子层的破坏和膜溶解,并根据不同类型的磷脂膜的结构变化,说明该抗菌肽与膜作用的机制为“毯式”模型(如图1所示)。
4展望
抗菌肽作为一种功能性添加剂用于分娩前后的母猪饲粮时,可以有效提高母猪的免疫机能和死产率、仔猪初生重和断奶重,显著降低哺乳期母猪产生的各种应激以及仔猪的腹泻率和死亡率,进而提高整体养殖效益。对于1000头母猪场而言,使用抗菌肽饲料添加剂可使年经济效益至少增加59.25万元。
但有关于抗菌肽的研究还有以下问题有待解决:
(1)抗菌肽是动植物免疫系统受到刺激后产生的物质。含量极微:虽然可利用化学方法合成,但成本较高,无法进行大规模的生产;(2)抗菌肽种类很多,有广谱的抗菌性:要发现一种生产成本低,使用效果好的抗菌肽添加剂,还需不断的研究探索;(3)抗菌肽种类丰富,抗菌机理各不相同,目前仍没有非常明确的、统一的理论;这急需研究者不断总结归纳抗菌肽的作用机制,加快抗菌肽应用的推广。
