小麦持久条锈病抗源品种89144(BJ144)芒性状遗传分析
欧巧明 崔文娟 李忠旺 王炜 陈琛 倪建福
摘要:分别以小麦持久条锈病抗源品种89144-2-3-11-2、89144-2-14-4-1-2为父本,感病小麦品种铭贤169为母本进行常规杂交,F1代种子单粒播种,在F2代成株期进行芒的分离遗传分析。结果表明,供试顶芒品系和全芒小麦杂交后,2组合F2代群体芒的性状分离均符合1∶2的理论比例,全芒对顶芒均为显性,且全芒受1对显性基因的控制。这是否说明小麦芒性或顶芒还存在隐性性状的可能,抑或与该小麦材料是外源DNA导入小麦的变异后代有关,需要进一步研究。
关键词:小麦;89144;条锈病;持久抗源;芒性状;遗传分析;隐性基因
中图分类号:S512.1? ? ? ?文献标志码:A? ? ? ? 文章编号:1001-1463(2020)10-0031-04
doi:10.3969/j.issn.1001-1463.2020.10.007
Abstract:89144-2-3-11-2 and 89144-2-14-4-1-2 were used as male parents, and Mingxian 169 was used as female parents for conventional hybridization. Seeds of F1 generation were sown in single seed, and the separation and genetic analysis of mangans were carried out at the plant growth stage of F2 generation. The results showed that after the hybridization of apical and total awn strains and total awn wheat, the traits of awn in F2 generation population of 2 combinations were all in accordance with the theoretical ratio of 1∶ 2, the awn was dominant to total awn, and the total awn was controlled by a pair of dominant genes. Whether this indicates the possibility of recessive traits in wheat awn or apical awn, or whether this wheat material is related to the mutant offspring of imported wheat DNA, needs further study.
Key words:Wheat;89144;Stripe rust;Persistent source resistance;Awn traits;Genetic analysis;Recessive gene
芒作为小麦穗器官的组成部分,是小麦重要的光合器官,其同化产物直接运往所着生的小穗,对籽粒充实起重要的作用。小麥芒性的表现分为无芒、顶芒和全芒3类,芒的有无、形状、长短及颜色等差异是品种易识别的性状特性之一。前人研究发现,小麦芒性性状具有提高光合面积和光合效率、固定CO2[1 ],提高小麦产量的作用,是一个对产量有直接影响的重要性状[2 - 3 ],体现在有芒的品种比无芒品种有更高的产量(通常增产10%以上)[4 - 5 ]。芒的光合产物对穗重的贡献可超过35%,甚至有芒品种穗对粒重的贡献大于旗叶[5 ]。有报道称。小麦芒的碳交换率约是整个穗部的40%~ 80%[6 ],其同化产物99%运入着生该芒的小穗[7 ]。但Weyhrich等[8 ]却认为无芒小麦比有芒小麦千粒重大。因此,研究小麦芒的特性及遗传规律对小麦育种和栽培具有重要意义。
国内外对于小麦芒性相关基因的研究普遍认为,无芒是显性性状[9 ],有多个基因决定芒的发育,不同基因组合决定不同类型的芒性[10 ],多数已被定位于小麦不同染色体 上[3,11 - 12 ]。王彦梅等[9 ]证实2A染色体有促进芒生长的隐形基因。Catkins A E [10 ]对小麦芒的遗传研究后指出,决定芒的发育和类型的5~6个主要基因中,分别位于5A、6B染色体长臂上的B1和B2基因须全部重合才能表现长芒[13 ],其中基因B1、B2、B3为芒抑制基因,B1作用最强,即使处于杂合态也表现无芒;B2次之,B3最弱,A基因促进芒生长[3,12 ]。Ahmad M[14 ]研究发现,5A、1B、3B、4B、5B、6B染色体上均具有促进芒发育的基因,而2A、6A、2B染色体上均具有抑制芒发育的基因,而影响芒长度的基因位于2A、3A、4A、1B、2B、3B、5B、2D、3D、6D和7D染色体上,对芒有抑制作用的基因位于5A、4B、5B、6B和1D染色体上[12,15 ]。Suneson[16 ]发现,7B染色体对芒表现有促进效应。目前,因不同研究者分类方法的不同及表现型相似基因型不同等原因,关于芒性状遗传的研究尚缺少一致意见。
目前国内外在小麦育种方面多倾向于有芒类型的选择和培育,对小麦芒形态及其特征研究方面开展的相对较多,但在其遗传规律方面相关报道较少。基于此,我们于2008 — 2010年开展了相关研究,现将结果报道如下。
1? ?材料与方法
1.1? ?材料
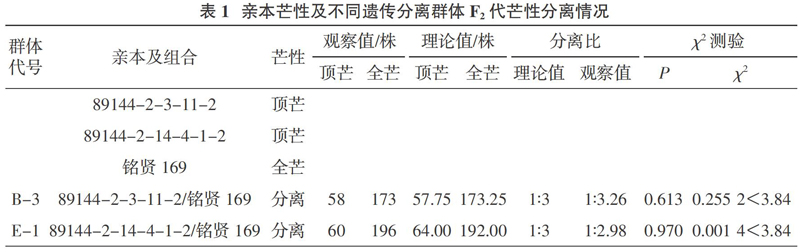
供试材料为小麦持久条锈病抗源品种89144-2-3-11-2、89144-2-14-4-1-2(甘肃省农业科学院生物技术研究所提供)和感病小麦品种铭贤169(西北农林科技大学提供)。其中持久条锈病抗源小麦品种89144-2-3-11-2、89144-2-14-4-1-2为甘肃省农业科学院生物技术研究所采用花粉管通道外源DNA导入技术将强抗逆作物高粱DNA导入感病受体小麦品种甘麦8号后,经连续选择变异系选育而成的,具有很好的持久条锈病抗性,其芒的特性见表1。
1.2? ?试验方法
2011年5月下旬以89144-2-3-11-2、89144-2-14-4-1-2为父本,铭贤169为母本杂交,2012年3月将杂交所获种子点播于甘肃省农业科学院生物技术研究所兰州试验点,2012年7月获得F1代种子。2012年10月将亲本及杂交F1代材料分别按组合单粒播种于温室,并于2013年4月中旬按杜斌[3 ]报道的芒性分类(即无芒、顶芒、全芒),逐株记载各杂交分离群体F2代芒的类型,进行芒的分离分析,用卡方进行适应性测验。
2? ?结果与分析
试验结果(表1)表明,89144-2-3-11-2/铭贤169组合F2代的长芒植株为173株,顶芒植株为58株,经卡方测验,符合1∶3的理论分离比例( χ 2(3∶1)=0.255 2 < 3.84),说明持久条锈病抗源89144-2-3-11-2芒的性状分离符合孟德尔规律,全芒对顶芒为显性,且全芒受1对显性基因的控制。89144- 2-14-4-1-2/铭贤169组合F2代的长芒植株为196株,顶芒植株为60株,经卡方测验,符合1∶3的理论分离比例(χ2(3∶1)= 0.001 4 < 3.84),说明持久条锈病抗源89144-2-14-4- 1-2芒的性状分离符合孟德尔规律,全芒对顶芒为显性,且全芒受1对显性基因的控制。
3? ?小结与讨论
小麦的芒不仅是小麦本身固有的特征,而且是影响小麦产量高低的重要上部器官。目前国内外对于小麦芒性相关基因研究普遍认为,小麦无芒对长芒是显性性状[9 ],有多个基因决定芒的发育,不同基因组合决定不同类型的芒性[10 ]。
本研究采用常规杂交方法,对2份持久条锈病小麦抗源材料的F2代分离群体的芒性进行遗传分析表明,长芒对顶芒为显性,且长芒受1对显性基因的控制,这与黄瑾 等[2 ]、杜斌[3 ]、骆惠生等[17 ]报道的小麦无芒对长芒是显性性状的结果不同。这是否说明小麦芒性或顶芒还存在隐形性状的可能,还有待进一步研究。
另外,供试小麦亲本89144是强抗逆作物高粱基因组DNA导入感病受体甘麦8号后选育的具有持久条锈病抗性的优良品系[18 - 20 ], SSR、RAPD等分子标记分析显示,供体高粱DNA可能已整合进受体基因组并得以表达[18 - 19 ]。抗锈机理分析显示,89144 在条锈菌侵染后表现出系统获得性抗性[21 - 22 ],HMW-GS的Glu-D1位点等位基因亦发生突变,由受体的2+12亚基变为5+10亚基[23 ],是优质且持久抗条锈病的优良种质资源[18 ] ,并在多年育种实践中作为抗锈亲本利用,与多个核心育种亲本具有良好的配合力。据我们推测,这种外源DNA导入后的性状变异和改良可归结为生物诱变所致和某种外源基因的定向转移所致[19 - 20 ]。本研究中小麦长芒对顶芒为显性的结论是否也与该小麦材料的特殊性有关,抑或是与外源DNA导入后的变异有关,也值得进一步跟踪研究。
参考文献:
[1] JOHNSON R R,WILLMER C M,MOSS D N. Role of awns in photosynthesis,respiration, and transpiration of barley spikes[J].? Crop Sci.,1975,15:217-221.
[2] 黃 瑾,骆惠生,张 勃,等.? 普通小麦芒的遗传分析[J]. 甘肃农业科技,2011,2:11-12.
[3] 杜 斌.? 小麦芒长抑制基因B1近等基因系的鉴定及遗传分析[D].? 泰安:山东农业大学,2010.
[4] ENANS L T,RAWSON H M. Photosynthesis and respiration by the fag leaf and components of the ear during grain development in Wheat[J].? Aust. Biol. Sci.,1957,23:245-254.
[5] 杨汉森,王世敬.? 春小麦籽粒增重过程及营养器官对于粒重的贡献[J].? 宁夏农林科技, 1987(1):16-20.
[6] BLUM A.? Photosynthesis and transpiration leaves and ears of wheat and barley varieties[J].? Exp. Bot.,1985,36:432-440.
[7] OLUGBEMI L B.? Distribution of carbon-14 assimilated by wheat awns[J].? Ann. Appll. Biol.,1978,90:111-114.
[8] WEYHRICH,R A,CARVER B E,SMITH E L.? Effects of awn suppression on grain yield and agronomic traits in hard red winter Wheat[J].? Crop Sci.,1994,34:965-969.
[9] 王彦梅,安调过,王志国,等. “高优503”小麦芒基因染色体定位[J].? 生态农业研究, 2000,8(4):31-33.
[10] Catkins A E,Ellerton S. The inheritance of glume shape in triticum[J].? Grnft,1940,40:243-270.
[11] 金善宝.? 中国小麦学[M].? 北京:中国农业出版社,1996:11.
[12] 薛 秀.? 小麦染色体工程与育种[M].? 石家庄:河北科学技术出版社,1993:43.
[13] KOSUGE K,WATANABE N,KUBOYAMA T,et a1. Cytological and microsatellite mapping of mutant genes for spherical grain and compact spikes in durum wheat[J].? Euphyfica,2008,159:289-296.
[14] AHMAD M.? Utilization of Fl monosomics for genetic analyses involving awn expression, glume color,seed setting,and seed abortion in crosses of tetraploid and hexaploid wheats[J].? Theoretical and Applied Genetics,1975,46:1-5.
[15] BOZZINI A,GIORGI B.? Genetic analysis of tetraploid and hexaploid wheat by utilization of monopentaploid hybrids[J].? Theoretical and Applied Genetics,1971,4(2):67-74.
[16] SUNESON C A, BAYLES B B, FIFIELD C C. Effect of awns on yield and market qualities of wheat[J].? US Dept. of Agriculture,1948,783:8.
[17] 骆惠生,曹世勤,贾秋珍,等.? 农家品种老芒麦抗条锈基因遗传分析[J].? 甘肃农业科技,2007(7):10-13.
[18] 倪建福,欧巧明,令利军,等.? 小麦抗条锈新品系89144抗锈机理研究[J].? 植物保护,2006,32(1):30-34.
[19] 欧巧明,崔文娟,王 炜,等.? 花粉管通道法导入高粱DNA创造优良小麦新品系的分子聚合育种[J].? 干旱地区农业研究,2013,31(2):6-12.
[20] 欧巧明,倪建福,崔文娟,等.? 高粱DNA导入引起小麦HMW-GS的变异及其品质改良和变异机理分析[J].? 中国粮油学报,2011,26(1):15-19.
[21] 裴新梧,崔凯荣,孔英珍,等.? 导入高梁DNA选育丰产、抗逆小麦新品系及其RAPD分子验证[J]. 兰州大学学报(自然科学版), 1999, 35(2):130-135.
[22] 邢更妹,王 蒂,崔凯荣,等.? 抗锈小麦品系89144对锈菌侵染和伤害反應的比较[J].? 中国农业科学,2003,36(1):44-48.
[23] 倪建福,欧巧明,庞斌双,等.? 高粱总DNA导入春小麦新品系高分子量麦谷蛋白亚基的变化[J].? 兰州大学学报(自然科学版),2005,41(6):47-49.
(本文责编:郑立龙)
