低温胁迫下桃子房和幼果的过冷却点及生理响应
王晨冰 王发林 万信 赵秀梅 牛茹萱


摘要:以17年生北京7号桃树枝条为试材,采用水培法,利用能准确模拟自然界霜夜降温过程的人工霜箱,测定桃盛花期花器、幼果的过冷却点、结冰点;同时对桃花器及幼果进行不同低温(15、0、-1、-2、-3 ℃)处理,测定桃花器渗透调节物质可溶性蛋白、脯氨酸、丙二醛(MDA)含量。结果表明,盛花期的花器过冷却点和结冰点均低于幼果,低温胁迫下渗透调节MDA含量随温度下降而增加,温度越低,MDA 含量上升趋势越明显,表现出一种应激反应;可溶性蛋白和脯氨酸含量均呈先升后降的趋势,表现为适应状。
关键词:桃;低温胁迫;过冷却点;丙二醛;可溶性蛋白;脯氨酸
中图分类号:S662.1? ? ? ?文献标志码:A? ? ? ? 文章编号:1001-1463(2020)09-0018-05
doi:10.3969/j.issn.1001-1463.2020.09.006
Abstract:Using 17-year old Beijing 7 peach tree branches as test material. The artificial frost box, which can accurately simulate the natural frost night cooling process and hydroponics was used to determine the over-cooling point and freezing point of the organs and young fruit in the flowering period. At the same time, peach blossom organ and young fruit were treated at different low temperatures(15 ℃, 0 ℃, -1 ℃, -2 ℃, -3 ℃) to determine the contents of soluble protein, proline and malondialdehyde(MDA) content. The results showed that the overcooling point and freezing point of flower tube were lower than young fruit in flowering stage, and the MDA content in osmotic regulation increased with the decrease of temperature under low temperature stress, the lower the temperature, the more obvious the rising trend of MDA content, which shows a stress response; The contents of soluble protein and proline increased first and then decreased, which was adaptive.
Key words:Peach;Low temperature stress;Supercooling point;Malondialdehyde;Soluble protein;Proline
秦安縣是甘肃省天水市桃栽培集中产区,桃园以浅山旱地为主,所产“秦安蜜桃”色艳味美,深受省内外消费者的喜爱。但是,由于当地山地地形复杂,区域性小气候特征显著,春季晚霜冻冻害发生频率极高。据2000 — 2010年期间完整的气象灾害调查资料显示,天水市春季晚霜冻冻害发生频率达60%左右,晚霜冻害已成为该地区主要农业气象灾害 [1 ]。晚霜冻害导致桃果大面积减产,甚至绝收,给生产带来巨大损失。因此,深入研究桃早期抗寒机理,对于选育优质、抗寒品种以及预防晚霜危害具有理论指导意义。有关桃霜冻害机理的研究已取得一定进展,如霜冻发生后对不同扁桃品种抗寒性的研究[2 ],白粉桃低温胁迫下花器的生理效应[3 ],低温胁迫对扁桃枝条细胞膜系统和渗透调节物质的影响[4 ]。关于桃花不同发育时期花器官、幼果抗霜的临界温度及低温对渗透物质调控未见系统化研究。我们利用人工模拟霜箱,以17年生北京7号桃不同花器官、幼果为试材,模拟天水浅山区自然界霜夜降温全过程,研究桃花器官、幼果的过冷却点和结冰点及低温条件下渗透调节物的反应,为研究桃花期霜冻机理及生产中预防晚霜冻害提供参考。
1? ?材料和方法
1.1? ?试验地点与供试材料
试验于2013年2月至2017年5月连续4 a在甘肃省天水市秦安县兴国镇柴家山进行。该地平均年降水量507.3 mm,年蒸发量1 423 mm;年平均气温10.0~11.4 ℃,活动积温3 382.2 ℃,日照时数2 208.1 h,无霜期176 d。试验地为黄绵土。
供试材料为17年生北京7号桃,行株距4 m×3 m,无灌溉条件,果园管理水平一般。于每年桃花花芽露红期采集带花枝条,在自然环境下水培,盛花后7 d采集桃枝条进行水培。
1.2? ?测试项目与方法
1.2.1? ? 桃花过冷却点的测定? ? 选择盛花期的花器测定过冷却点、结冰点,选择盛花后 9 d的外果皮作为幼果测定部位。提前将供试桃枝条用水培法置于人工试验霜箱内,利用 PT-100 型热电偶温度传感器探头测定桃花器及幼果果皮。人工模拟天水地区自然霜夜降温过程,设置降温速度为1 ℃/0.5 h。试验共设处理-10~-2 ℃共11个温度层次,温度降到设定值时并且保持 0.5 h。然后模拟自然界相同季节晴天的升温过程,以1 ℃/0.5 h速度升温。当温度上升到 7 ℃ 以上后,取出试验材料置实验室水培,1 d后进行相关指标测定。
1.2.2? ? 渗透调节物测定? ? 用水培法将桃花培养至一半花盛开,幼果枝采集大田内盛花后7 d桃枝条再进行水培。将试验材料分批放入以 PID 调节方式控温和 FrosTem40 数据采集的 MSZ-2F 型模拟霜箱内,设15、0、-1、-2、-3 ℃共5个处理。根据秦安发生霜冻时温度变化情况,模拟霜箱试验温度先预冷至 10 ℃ 左右,然后以 6 ℃ /4 h速度降温至 4 ℃ 左右,再以 1 ℃ /0.5 h的 速度缓慢降温,直到设定温度,保持 0.5 h后,设置升温速度为1 ℃/0.5 h,最终温度升至室内温度。以自然常温(15 ℃)下培养的桃花为对照(CK)。
丙二醛(MDA)含量的测定采用硫代巴比妥比色法[5 ],单位为nmol/g·FW;可溶性蛋白含量测定采用考马斯亮蓝 G-250 染色? ? 法[6 ],单位为ng/g·FW;脯氨酸含量测定采用酸性茚三酮法[7 ],单位为ug/g·FW。
试验数据用SPSS 16.0软件进行方差分析,差异显著性分析采用Duncan新复极差法。
2? ?结果与分析
2.1? ?盛花期花器及幼果的过冷却点及结冰点
当植物体温度下降到 0 ℃以下时,植物器官组织内水分并不马上结冰而仍然保持液态,这种现象称之为过冷却状态(图1)。过冷却点的高低是判断植物抗寒性强弱的标志之一,过冷却点越低,抗寒性越强。研究表明,北京7号盛花期花器的过冷却点为-7.5~-4.5 ℃,幼果过冷却点为-5.5~ -3.5 ℃(表 1),说明随发育进程的推移,幼果的抗寒能力较花器弱。北京7号的桃花器、幼果均可以利用过冷却避免-3 ~0 ℃的结冰危害,但随着外界温度持续降低而低于植物组织自身能抵御的最低温度时,组织释放潜热,温度突然上升并开始结冰(结冰点),组织器官温度一旦冷却点跳跃上升至结冰点后便开始结冰,温度回落再升至室温后花器和幼果均发生褐变。表明花器和幼果不耐结冰,是依靠保持过冷却状态避免冰晶形成来抵御霜冻,过冷却点温度恰好是植物组织器官生理适应的低温下限。
2.2? ?低温胁迫对渗透调节物的影响
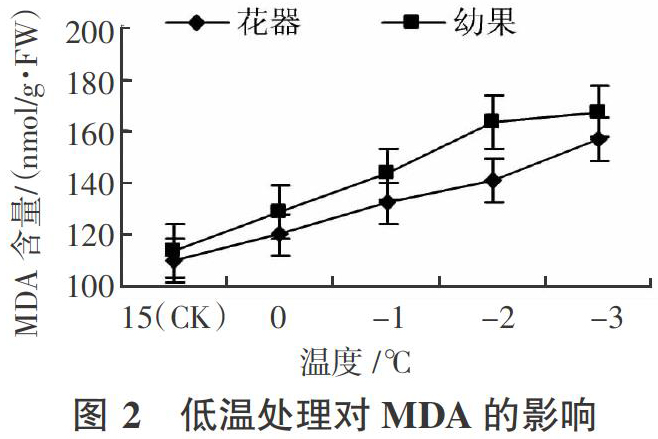
2.2.1? ?对 MDA 含量的影响? ? MDA是细胞膜脂过氧化的产物,也是膜系统受害的重要标志之一。由图 2 可知,花器和幼果中的MDA含量随温度下降而增加,MDA含量变化趋势是缓慢增长到急剧上升之后缓慢上升。花器和幼果MDA增加的幅度和临界温度不同,幼果的增加幅度较花器大,同时比花器提前出现缓慢增长期。花器在温度低于-2 ℃时增幅较大,花器与幼果在-2 ℃时MDA 含量分别较CK增加30.6 nmol/g·FW和29.1 nmol/g·FW。
2.2.2? ? 对可溶性蛋白含量的影响? ? 图 3 为低温处理下花器官及幼果可溶性蛋白含量变化。可以看出,从常温15 ℃降到0 ℃时,可溶性蛋白增长缓慢;当温度降到-1 ℃时,可溶性蛋白含量迅速升高。与CK相比,花器与幼果在-2 ℃时分别增加了 15.9 ng/g·FW和28.7 ng/g·FW。
2.2.3? ? 对脯氨酸含量的影响? ? 脯氨酸常被作为植物抗逆性的指标,是種膜保护物质,含量越高越有利于抗寒。低温条件下花器官及幼果脯氨酸含量的变化如图4所示,花器及幼果脯氨酸含量随着温度下降均呈上升趋势。与CK比较,在-2 ℃花器及幼果分别较对照增加16.1 μg/g·FW和17.2 μg/g·FW。
3? ?小结与讨论
西北地区晚霜冻冻害是制约北方落叶果树生产发展的一个难题[8 ]。我们采用能够准确模拟自然界霜夜降温过程的人工霜箱,研究了春季桃花和幼果发育期的过冷却点和结冰点温度的变化。研究发现,桃北京7号桃花器的过冷却点的变化范围在3 ℃以内,而幼果冷却点的变化范围在2 ℃以内。幼果过冷却点和结冰点均高于花器,表明幼果御寒能力较桃花器低,这与王飞等[9 ]在杏花上的研究结果一致,说明植物花器本身的抗寒性与其发育程度有关。
植物的耐低温机制相当复杂[10 ]。研究表明,经低温处理后花器及幼果MDA含量迅速增加,说明细胞膜对低温伤害也是一个逐步适应直至最后破坏的过程,也是一种应激反应过程。MDA作为膜脂过氧化的最终产物,能够抑制细胞保护酶活性和降低抗氧化物的含量,从而加剧了膜脂过氧化性[11? - 12 ],这一结果与杨春祥等[13 ]低温胁迫对油桃花器生理变化的研究结果一致。低温可以引起植物细胞蛋白质的变化主要表现在可溶性蛋白和相关酶的协同变化结果导致产生抗寒性蛋白。可溶性蛋白具有较强的亲水性,它能明显增强细胞的持水力,而可溶性蛋白含量的增加可以束缚更多的水分,减少低温条件下原生质因结冰而受伤害致死的机会[14 ],从而提高植物体抗寒性。低温胁迫过程中,桃花器和幼果的可溶性蛋白含量随着温度降低逐渐升高,这可能是机体对低温环境逐步适应的表现。脯氨酸是植物体内的重要渗透调节物质,在低温条件下,脯氨酸大量积累,提高了细胞液的浓度,维持细胞的膨压,降低冰点,防止原生质过度脱水[15 ]。本研究中桃花器及幼果的MDA、可溶性蛋白及脯氨酸含量升高,与花器和幼果过冷却点相对应。
桃花的冻害程度不仅取决于栽培品种、地形、土壤及管理水平的内在差异,低温强度、低温持续时间、环境湿度、风速等外界环境条件的影响也极为显著[16 ]。相关研究表明,桃树花芽在萌动后花蕾变色期的受冻温度为-6.6~-1.7 ℃,开花和幼果期的受冻温度为-2.0~-1.0、-1.1 ℃[17 ]。本试验的环境条件与桃园环境下自然花器的抗低温能力及风速、水汽等条件存在差异,冻害指标研究仅建立在统计学基础上,但结果与当地冻害年份霜冻灾害实况基本一致,在实际生产中具有一定的参考价值。
参考文献:
[1] 赵秀梅,牛茹萱,张? ?帆,等.? 8个桃品种在兰州地区的抗寒性鉴定[J].? 甘肃农业科技,2019(12):43-46.
[2] 王? ?森,谢碧霞,杜红岩,等.? 美国扁桃花器官的抗寒性[J].? 经济林研究,2007,25(2):19-22.
[3] 倪鼎文,李? ?唯,杜志钊,等.? 外源SA对白粉桃低温胁迫下花器的生理效应[J].? 甘肃农业大学学报,2007,42(6):53-56.
[4] 龚? ?鹏,李秀霞,李增萍,等.? 低温胁迫对扁桃枝条细胞膜系统和渗透调节物质的影响[J].? 新疆农业科学,2007(5):561-566.
[5] 白宝璋,郭守华,杨晓玲,等.? 植物生理生化:下[M].? 北京:中国农业科技出版社, 2003:244-246.
[6] 邹? ?琦.? 植物生理学实验指导[M].? 北京:中国农业出版社,2000:57-58.
[7] 邹? ?琦.? 植物生理学实验指导[M].? 北京:中国农业出版社,2003:166-167.
[8] 杨建民,周怀军,王文凤.? 果树霜冻害研究进展[J].? 河北农业大学学报,2000,23(3):54-58.
[9] 王? ?飞,陈登文,李嘉瑞,等.? 杏花及幼果的抗寒性研究[J].? 西北植物学报,1995,15(2):133-137.
[10] 何维勋,冯玉香,夏满强.? 解冻速率对作物霜冻害的影响[J].? 应用气象学报,1993,4(4): 440-445.
[11] GAO AINOG,JIANG SHU RONG,ZHAO XIWEN,et al. Study on cold hardiness of apple cultivars[J].? Journal of Fruit Science, 2000,17(1):17-21.
[12] PRASAD T K. Role of catalase in inducing chilling tolerance in pre-e-mergent maize seedlings[J].? Plant Physiol.,1997,114(4):1369-1376.
[13] 楊春祥,李宪利,高东升,等.? 低温胁迫对油桃花器官膜脂过氧化和保护酶活性的影响[J].? 果树学报,2005,22(1):69-71.
[14] 王? ?毅,杨宏福,李树德,等.? 园艺植物冷害和抗冷性的研究[J].? 园艺学报,1994,21(3):239-244.
[15] 沙? ?伟,刘焕婷,谭大海,等.? 低温胁迫对扎龙芦苇SOD、POD 活性和可溶性蛋白含量的影响[J].? 齐齐哈尔大学学报,2008,24(2):1-4.
[16] 周蕴薇,聂绍荃.? 翠南报春抗寒生理生态学研究[D].? 哈尔滨:东北林业大学,2001.
[17] 许彦平,姚晓红,刘晓强,等.? 近30 a甘肃天水气候资源变化对杏产量影响评估[J].? 干旱区地理,2015,38(4):684-691.
(本文责编:陈? ? 珩)
