
摘要:自噬是对受损的蛋白质和细胞器的消除主要代谢途径,是目前公认的一种抗衰老的过程和维持细胞内环境稳定的基础作用,同时自噬也是适应生物力学刺激的一种响应机制。通过自噬在骨组织细胞和骨代谢疾病中的重要作用,提出运动可调节骨组织细胞的自噬。方法:回顾近年来有關在不同骨细胞类型即成骨细胞、破骨细胞和骨细胞的自噬作用的文献资料法。结果:通过近年文献的综述,发现骨组织细胞自噬与骨代谢、骨相关疾病有着重要的联系。结论:自噬在骨生理学、骨代谢中存在重要作用,运动作为调节自噬的一种重要方式,可能调节不同骨细胞类型自噬,进而影响骨代谢。
关键词:运动;自噬;骨细胞
中图分类号:G 804.2 文章编号:1009-783X(2017)02-0171-06 文献标志码:A
运动调节骨代谢的作用在国内外已有很多研究,运动对骨代谢的调节是通过多种途径整合的过程,例如运动调节骨组织细胞信号通路、激素分泌,从而调节骨代谢等。现在发现骨组织细胞自噬作为一种重要的途径,存在饥饿和应激状态下,对细胞内组分进行自我消化,获取养分、维持能量平衡,调节骨代谢。研究发现,骨组织细胞自噬的发生在一定程度上有利于细胞存活。细胞质靶向降解物质包括长寿蛋白,聚集体和受损的细胞器。双膜囊泡的自噬体与溶酶体融合形成自噬溶酶体后,内容物降解和循环利用。基础自噬发生在所有细胞中较低的水平,是一种蛋白质和细胞器的质量控制机制,维持正常的细胞内环境的稳定。此外,当遇到各种压力如饥饿、氧化应激、缺氧或感染时自噬上调,类似一种保护机制。骨是一个复杂的器官,骨的重建贯穿人的一生,有一个严密的控制机制,主要由3种细胞类型执行:成骨细胞(Osteoblast,OB)、破骨细胞(Osteoclast,OC)、骨细胞(Osteocyte,OST)。研究发现这3种骨组织细胞均存在自噬,运动作为调节自噬重要的方式是否调控骨细胞自噬进而诃节骨代谢?本文对近年来关于骨组织细胞自噬的文献进行综述,旨在为运动调节骨组织细胞自噬进而调节骨代谢进行展望,这将成为运动调节骨代谢的一个新课题。
1.自噬的机制
哺乳动物雷帕霉素靶蛋白(mammalian target of rapamytin,roTOR),--fO高度保守的丝氨酸/苏氨酸激酶,是mTORCl复合物的一部分,包括Raptor和mLST8蛋白,作为营养和能量的传感器,是调控自噬的主要途径。
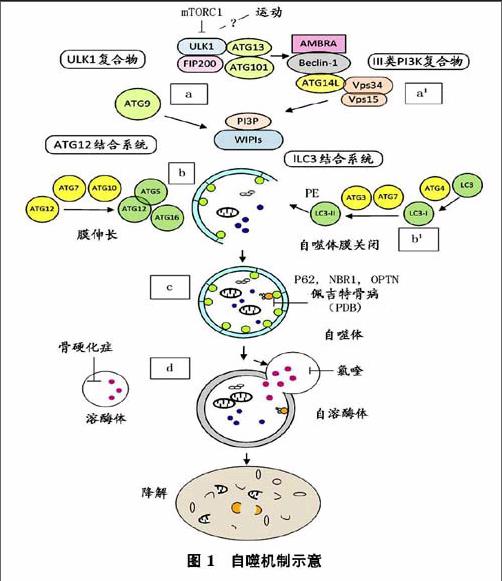
自噬体的形成是一个多步骤的过程(如图1所示),包括超过15个自噬相关蛋白(ATG)分层招募前自噬体结构(PAS),a在应对不同刺激如mTORCl失活,细胞自噬是通过ULKl复合物和Ⅲ类P13K复合物启动。由于局部PBP形成,与ATG9提供部分膜相关,形成一个吞噬泡。b该ATGl2共轭体系与LC3的共轭体系一样,都涉及隔离膜的伸长和关闭。自噬体融合溶酶体形成一个完整自噬溶酶体,通过水解酶降解的内容材料,释放在细胞质中代谢产物作为能源。a由ULK-1/2,ATG-13,ATG-101和FIP200组成(ULK)复合物介导启动自噬。在营养充足的条件,mTORCl通过ULK-1/2、ATGl3的磷酸化负向调节ULK复合物。在运动和饥饿刺激下,可能导致mTORCl失活,激活ULK-1,ATGl3和FIP200磷酸化,诱导自噬发生。a'ULKl的磷酸化触发多蛋白复合物的易位(Ⅲ类P13K复合体I),含Beclin-1(也被称为ATG6)、AMBRA、ATG14L、Vpsl5和Ⅲ类磷酸激酶(P13K CⅢ,也称Vps34),从细胞骨架结构到细胞自噬前体到初级自噬体的形成。磷脂酰肌醇-3(P13P),由VPS34活动产生,约束W1PI(WD重复蛋白与磷酸肌醇相互作用)效应器和ATG9一起调节自噬体形成的最初阶段,ATG9在不同区域循环,被认为是一个膜延伸脂质载体。
延伸和闭合过程需要2个泛素样蛋白复合物及相应的共轭机制和几种SNARE蛋白。b首先包括ATG7和ATGl0酶,来调节泛素样ATGl2到ATG5的共价结合ATGl2-ATG5。ATGl2-ATG5结合后再结合到ATGl6形成ATGl2-ATGS/ATGl6,是自噬体生物合成必不可少的过程,然后结合到PAS第2类泛素样系统。bl第2类泛素系统允许磷脂酰乙醇胺(PE)共轭到微管结合的轻链3(LC3),与酵母ATG8同源。初期的LC3是ATG4首先产生的,由ATG7活化,转移到ATG3缀合酶,再结合到PE。LC2-PE(LC3Ⅱ)作为自噬的标记,目前是在自噬体上完整的膜蛋白。
自噬作用一直被认为是一种非选择性的过程,但几个选择性自噬形式已经被确定。自噬受体一般出现一个泛素结合结构域允许破坏性的泛素化结构相互作用,一个特定的氨基酸序列为LC3相互作用区(LIR序)与ATG8蛋白家族成员结合。除了其降解功能,最近在OC观察到蛋白质分泌自噬,此外,在生物力感应细胞还发现自噬有胞吐作用。
2.骨组织细胞中的自噬调节器
通过对近年来文献的查阅整理,发现几个有利于骨骼健康自噬诱导剂,运动可能诱导调节以下诱导剂,调节骨组织细胞的自噬功能从而调节骨代谢,见表1。
3.骨组织细胞自噬
越来越多的文献表明,骨细胞自噬功能的缺失可能导致骨细胞功能障碍,以下就最近关于间充质干细胞(mesenchyma stem cell,MSCs)、OB、OST和破骨细胞(osteoclast,OC)自噬作用的研究结果进行综述。
3.1自噬与MSC
MSCs具有干细胞特性,即自我更新和多向分化潜能,使他们能够分化成多种细胞包括成骨细胞、软骨细胞和脂肪细胞。Song等表明,自噬诱导MSC氧化应激可导致生存反应。另外,自噬还被证明参与维持间充质干细胞干性。最近,nuschke等观察到在未分化的间充质干细胞自噬通量停止,产生大量的未降解的自噬小泡积累,在向OB分化早期,这些自噬小体快速产生能源物质供MSX消耗。而研究推断,运动可能通过AMPK先抑制mTOR调节的自噬和后激活Akt/mTOR信号轴,调控人骨髓问充质干细胞向成骨细胞分化。
3.2自噬与UB和UST
OB来源于间充质细胞,参与骨的形成和矿化。成骨细胞在骨形成过程中,一些成骨细胞被包埋在自己的基质,分化成“骨细胞”,但大多数最终凋亡。采用成骨细胞系、颅骨或BMSC得到的OB表明,OB分化和矿化过程中自噬功能增加,而运动促进OB分化可能与运动过程中自噬功能增加有关。使用透射电子显微镜分析小鼠OB,观察双层膜自噬囊泡的存在,表明细胞内矿化机制受自噬调节。通过自噬抑制OB分化和矿化,自噬作用在OB进一步证实。对骨髓基质干细胞用3-1VIA和氯喹晚期治疗(在分化10 d)会引起碱性磷酸酶阳性细胞数量和大小减少,导致在21 d矿化比对照培养组显著减少…。这些治疗对早期的OB分化无明显影响,表明OB终末分化需要自噬,而运动可能增加终末端的自噬功能从而促进OB终末端分化与矿化。d同样,巴弗洛霉素,氯奎因和氯化铵,以及shRNA介导降低自噬必需基因LC3,阻断人牙髓间充质干细胞,成骨分化hDP-MSC。最后,siATG7和siBECNl都大幅降低了大鼠成骨细胞系的矿化效率,因此,除了间接作用于OB分化,自噬似乎通过细胞外的分泌直接参与矿化过程。骨细胞代表成骨细胞系的终末分化阶段,被嵌入在矿化骨基质,因为它们的寿命长,位置固定,这些细胞在适应紧张的环境时,為存活将高度依赖自噬。研究发现,在小鼠和人的皮质骨骨细胞间断分布的LC3,引起高水平的基底细胞自噬。此外,OST在饥饿和缺氧条件下诱导自噬,糖皮质激素也诱导自噬,从而保持OST活力。OB和OST是敏感细胞,调节应力加载后骨适应性反应,运动力刺激可能翻译成结构级联性生化变化,有效刺激骨细胞自噬,从而提高骨强度,抑制随着年龄增长的骨损失。在体内遇到机械压力(0.2 kPa)时,强烈诱导OB自噬,且短暂和不依赖mTOR的。自噬在控制骨骼平衡的OB和OST功能上发挥着核心作用,这可能为运动调控骨代谢研究提供了新的方向。
对体内构建成骨细胞自噬缺乏的小鼠模型如下;首先,OSX-Cre小鼠骨髓基质细胞表达Cre重组酶成骨细胞向的骨原细胞与FIP200floxed小鼠杂交发现,突变的小鼠表现出骨小梁体积减少的骨质疏松表型(BV/TV)和显著的骨皮质厚度减少。组织形态学分析表明突变的动物OB数量和表面积与对照组相比减少,骨形成减少,OC数量和表面不变。Liu等使用骨髓基质细胞研究还发现,这些突变小鼠OB标记物的表达和矿化减少。总之,这些结果表明自噬在OB增殖具有重要作用。
其次,使用COLlA Cre小鼠在OB表达Cre重组酶,和ATG5 floxed小鼠杂交,得到OB特定自噬缺陷小鼠,然后对九月龄雌性和雄性小鼠股骨组织形态学和mlcroCT分析显示,突变小鼠的骨小梁体积减少,伴随着骨小梁宽度和数量降低。这种影响在雌性小鼠比雄性更明显。在两性的突变小鼠OB著减少,0{3周长增加,雌、雄突变小鼠OB/OC与对照组相比显著减少。此外,9个月大突变小鼠与对照组相比矿物沉积率下降50%。采用体外分析发现自噬缺陷OB呈现氧化应激增加和NF-vJ3受体活化因子(receptor activator of nuclear factor Bligand,RANKL)的分泌,有利于OC生成。研究得出,运动缺乏可能降低自噬使其矿化能力降低,引发OB和012之间的不平衡导致低骨量表型。
同样,利用COLlA-Cre小鼠在OB表达Cre重组酶与P62 floxed小鼠杂交生成的OB特殊P62缺陷小鼠表现出OB分化减少和骨量降低,在这些突变小鼠中观察发现骨小梁数量及骨细胞数显著减少。此外,OB缺乏P62导致骨髓内环境损失。自噬在0ST作用是使用Dmpl-Cre在OST表达ere和一些成熟的OB ATG7iloxed小鼠杂交,在小鼠6周测量BMD显示出雄性条件敲除小鼠股骨的骨密度略有下降,但没有改变脊柱和全身骨密度。检测到的雌性小鼠在这个年龄在任何位置BIvlD均没有变化。相比之下,在6个月的年龄条件基因敲除雄性和雌性小鼠在所有骨骼部位骨量与对照组相比下降。在骨形态计量学被证实OB和OC数目和面积减少以及脊柱骨形成率(BFR)降低,矿化沉积率(MAR)没有变化、股骨骨小梁体积减少,股骨皮质厚度减少。总之,这些数据表明,OST自噬功能障碍,类似骨骼老化骨重建率降低,运动对OST自噬的调节有待进一步验证。
3.3自噬与OC
OC通过OB产生M-CSF和RANKL触发融合成巨大、多核细胞,这些细胞负责骨吸收。最近发现自噬参与上调Beclin-1、MCP-1(单核细胞趋化蛋白1)和RANKL的表达促进OC分化。TRAF3是通过自噬降解负调控RANKL诱导OC形成的调控因子。MCP-1是一个CC趋化因子,诱导单核细胞分化成TRAP和CTSK阳性的OC前体细胞。这种效应被证明是在人外周血单核细胞通过锌指蛋白诱导的,称为MCPIP。wang等的研究表明通过MCPIP诱导的氧化应激,Beclin-1表达上调和自噬调节MCP-1诱导OC前体细胞分化。这个过程中ATG7下降抑制(X3标记TRAP、组织蛋白酶K(CathepslnK,CTSK)的表达,表明自噬参与OC前体细胞的分化。也有学者用骨髓巨噬细胞(BMM)在RANKL诱导破骨细胞产生过程中发现,Beclin-1不依赖自噬作用,而是通过诱导活性氧及主要基因NFATcl的表达。此外,如缺氧或微重力的诱导下,破骨细胞受自噬调节而增加,例如耐力运动缺氧或失重状态下可能通过自噬而调节破骨细胞增加,从而导致骨量丢失的。此外,TRAF3抑制RANKL,降低了RANKL对骨髓来源的破骨细胞前体的形成,这可能是运动过度或骨废用状态下骨量下降的又一重要原因。
这些数据表明,自噬参与OC的分化。OC的分化,OC皱褶缘生成和骨吸收需要ATG5,ATG7,ATG4B和LC3相关蛋白。ATG5与LC3-Ⅱ靶向皱褶缘,LC3-Ⅱ这个膜蛋白能促进分泌骨吸收需要的溶酶体。另外,小鼠OC里ATG5特异性敲除导致卵巢切除小鼠骨量增加,骨丢失减少,发现运动可能是抑制OC自噬从而抵抗骨质疏松症。下面列举骨组织细胞自噬与骨代谢疾病。
4.自噬与骨疾病
4.1佩吉特骨病
佩吉特骨病(PDB)是骨吸收增加和骨形成紊乱的非连续性病变。在细胞水平,PDB是破骨细胞过度活跃表现,例如体积、核数的增加。在SQSTMl/P62基因的泛素相关结构域突变已被证明参与了大部分的PDB,此外,它作为自噬受体的作用,P62参与NF-rB信号通路,调控OC的形成和凋亡。虽然感染等其他环境因素也被认为是这个病理辅因子,但Daroszewska等研究证明,P62UBA域点突变足以引起小鼠poget's病。在这个模型中,与OC对照组相比,在巴弗洛霉素A1突变时LC3-Ⅱ上升,表明自噬体形成增加。最近,全基因组关联方法确定cOPTN基因变异是PDB的危险遺传因素。OPTN在各种生理过程包括NF-gB调控中是重要的一员,此外,P62、OPTN作为自噬受体参与蛋白质聚集体的清除;因此,除了在骨生物学的作用,这2个蛋白也是参与细胞自噬的2个关键因素。在PDB中清除聚合物缺陷和改变自噬的作用还在进一步研究中。
4.2石骨症
常染色体隐性遗传的石骨症(ARO),也被称为恶性婴儿骨硬化病,是一种遗传和表型异质性OC功能障碍为gc-K的疾病。ARO患者表现出骨量增加,形态正常OC由于皱褶缘形成的缺陷无法再进行骨吸收。观察出生后不久,如果对这种病不进行治疗通常会致命。一个具有纯合突变ARO小鼠模型TCIRGl基因小鼠编码V-ATPase下游的a3亚基被最近证明早期自噬缺陷,在突变0C,LC3Ⅱ表达减少P62表达增加。有趣的是,在ARO两突变基因,CLC7和OSTMl与神经退行性疾病和LC3一Ⅱ的表达增加相关,可能是由于通量扰动原因所致。
4.3骨质疏松
与年龄相关骨丢失的骨质疏松症,主要是影响年龄超过50岁以上的人的骨脆性综合症。骨衰老潜在的分子机制仍在继续研究,除了性激素的损失,氧化应激增加被认为是这个过程的一个关键因素。自噬被报道发生在一些与年龄相关器官如肝、骨骼肌、肾与胰岛细胞,现在还发现自噬也可能是与年龄相关的骨丢失的一个重要因素,随着年龄的增加骨细胞P62的表达和细胞凋亡也出现上升趋势,LC3-Ⅱ、BECNl、ULK-1表达下降;因此,氧化应激所致细胞自噬的下降,可能是参与年龄相关骨丢失因素之一,而运动参与自噬调控进而阻止衰老引起的骨量丢失。研究发现细胞自噬功能的下降是加重雌激素缺乏导致骨质流失的一个重要因素。最后还发现,通过对人类全基因组关联数据挖掘研究发现,自噬基因与人类高度和骨质疏松症是高度关联,运动尤其是不同的运动方式如何调节骨细胞自噬基因从而防治骨质疏松,有待我们进一步求证。
5.运动调控骨细胞自噬的研究展望
骨组织细胞自噬对骨骼健康至关重要,而运动与自噬又有着千丝万缕的联系。自噬出现在OB和OC分化,OB、OC是参与骨构建与重塑的重要的骨组织细胞,且OB的存活和功能需要自噬过程,如骨基质的矿化,同样几个自噬蛋白参与OC的骨吸收,而骨重建的长寿关键调节器OST,也高度依赖自噬生存。一些骨疾病,如骨质疏松症与自噬缺陷息息相关,而科学的运动可以提高骨量,有效预防骨质疏松,自噬作为调控骨组织细胞代谢重要的方式,可能与科学的运动方式存在一定内在联系。随着生物医学和运动科学领域认识的不断深入,将进一步揭示运动如何调控骨组织细胞自噬从而调节骨代谢a,成为运动科学领域研究运动对骨代谢生物学机制的新方向,希望为运动健骨在运动科学领域找到更多科学的理论解释。
- 基于受众视角的公众人物“人设”传播学解读
- 守护好报纸的质量“生命线”
- 《经典咏流传》融媒体创新传播模式探讨
- 新时代综艺节目导演的创新研究
- 《大众日报》区域观察报道特色浅析
- 新传播环境下的新闻立台
- “情景再现”在播音主持中的应用
- 场景传播兴起的动因及影响分析
- 传统媒体如何转变思维打造融媒体
- 新闻记者的策划意识及提升对策
- “互联网+传统媒体”媒介互通新探究
- 分析新媒体时代媒介传播面临的伦理困境
- 融媒体时代电视新闻编辑的创新意识和融合能力
- 新媒体环境下高校舆论引导探析
- 如何正确应对自媒体时代舆论风向的多变性
- 新媒体时代广播新闻发展探析
- 互联网背景下不同新闻媒体标题语言的异同
- 新时代民生新闻报道的发展要点
- 广播电视中播音主持语言表现的魅力
- 传播学视野下广播电视编导专业定位再思考
- 智能化监控系统在中波台的应用
- 广播电视固态发射机原理及维修策略
- 频率专业化与广播节目创新的思考
- 语言类广播节目的录制技巧
- 多媒体粒子光效在电视节目制作中的运用
- abhor
- abhorred
- abhorrence
- abhorrences
- abhorrent
- abhorrently
- abhorrent-of
- abhorrent-to
- abhorrer
- abhorrers
- abhorrible
- abhorring
- abhors
- abide
- abide-by
- abide by
- abide by sth
- abided
- abided-by
- abider
- abiders
- abides
- abides-by
- abiding
- abiding-by
- 七宝烧
- 七宝粥
- 七宝羹
- 七宝莲花
- 七宝鞭
- 七家茶
- 七尊八点
- 七尺
- 七尺之形,不如一尺之面
- 七尺之躯
- 七尺变六尺——脑袋搬家
- 七尺布拦腰剪——不三不四
- 七尺昂藏
- 七尺汉子六尺门
- 七尺牛
- 七尺长的被子——满展
- 七岁做和尚,八岁做道士——玩鬼出身
- 七岁八岁讨狗嫌
- 七岁看大,八岁看老
- 七巧
- 七巧巧
- 七巧板
- 七年之病,求三年之艾
- 七年战争
- 七庙