和文志 潘鹏举 杨玲
摘要:为进一步明确铁线莲属16个园艺品种之间的亲缘关系和遗传多样性,利用12条ISSR引物对16份材料进行PCR扩增,共获得117条清晰的条带,多态性条带占比为97.44%。利用引物UBC815和UBC817构建16个铁线莲品种的指纹图谱,可以有效地对铁线莲进行区分和鉴定。利用NTSYS软件计算出铁线莲品种间的遗传相似系数为 0.20~0.73,平均相似系数为0.51。通过UPGMA法进行聚类分析,在相似系数为0.53时,可将16份材料划分为4大类。结果表明,铁线莲品种之间的遗传关系较为复杂,利用ISSR分子标记技术可以从分子水平揭示16份材料的遗传多样性水平和亲缘关系。
关键词:铁线莲;品种;ISSR;遗传多样性;指纹图谱
中图分类号: Q756;S682.390.3? 文献标志码: A? 文章编号:1002-1302(2019)12-0079-04
铁线莲为毛茛科(Ranunculaceae)铁线莲属(Clematis L.)植物,现已知的铁线莲约有300种,广泛分布于世界各地。我国约有133种,主要分布在华中和西南地区[1]。铁线莲具有非常高的药用价值和观赏价值。该属植物不仅花型多样,有平盘形、十字形、钟形、长瓣形等,还兼具花期长、色彩丰富的特点,拥有“藤本皇后”的美誉。在园艺家和植物学家几百年的栽培和育种过程中,培育出了上千个各具特色的铁线莲园艺品种[2]。这些品种中,绝大部分都是由多个不同的种杂交而来,还有部分品种是由野生种驯化而来[3]。长时间的人工选择过程中,铁线莲品种的遗传背景和各品种之间的亲缘关系变得模糊不清,有些杂交品种已经找不到最初的杂交亲本,这给杂交育种亲本选择带来一定的困难。同时,铁线莲很多品种外观形态相似度高,导致铁线莲品种鉴定越来越难。由于受环境影响因素较大,传统形态学分类方法难以对铁线莲品种进行有效区分。因此,运用分子生物学手段对铁线莲品种遗传多样性和亲缘关系进行研究,可为铁线莲品种在分子水平的分类与鉴定提供依据。目前,在有关铁线莲品种遗传多样性研究上,前人多从形态学和细胞学[4-7]等方面开展了一些研究,利用分子生物学手段进行相关研究的报道较少。
简单重复间序列(inter-simple sequence repeat,ISSR)标记技术具有操作简单、稳定性好和多态性丰富等特点,广泛应用于植物品种鉴定、基因作图与定位、遗传多样性和系统发育等的研究。孙正海等建立了铁线莲ISSR-PCR的反应体系,并且进行反应体系优化和引物筛选,但未进一步研究该分子标记技术在铁线莲品种遗传多样性研究中的可行性与稳定性[8-9]。本研究利用ISSR分子标記技术从分子水平对铁线莲16个园艺品种遗传多样性进行探究,为ISSR分子标记技术在铁线莲品种分类鉴定、亲缘关系分析以及种质资源利用和新品种培育过程中亲本选择提供理论依据。
1 材料与方法
1.1 材料
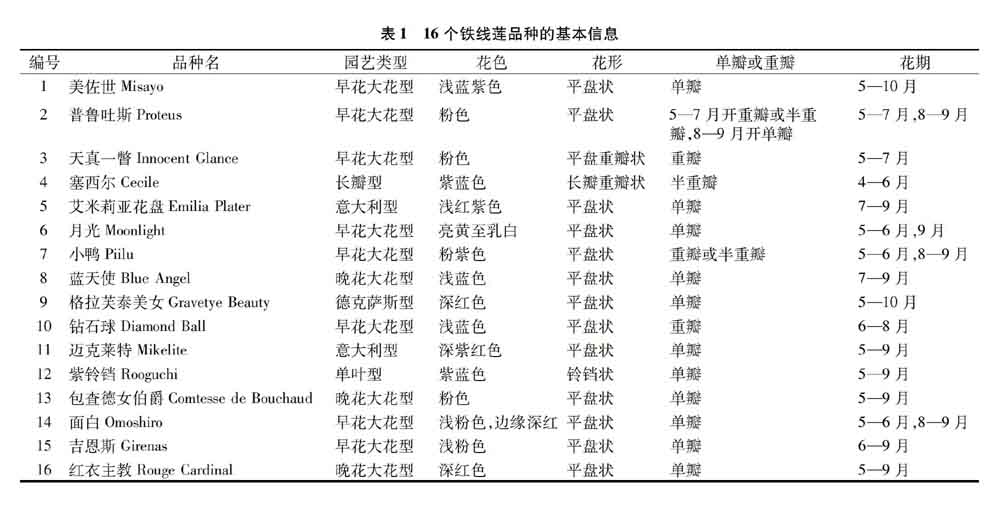
试验材料为2015年12月从国外引进的16个铁线莲园艺品种。试验地为西南林业大学一号教学楼前苗圃,位于云南省昆明市盘龙区,属于北亚热带低纬高原山地季风气候,年平均气温16.5 ℃,年均降水量1 450 mm,无霜期278 d。采集生长良好的植物幼嫩叶片,置于放入硅胶的自封袋当中,带回实验室置于-80 ℃冰箱保存备用。品种信息(来自世界园艺铁线莲协会)详见表1。
1.2 方法
1.2.1 DNA提取与检测 采用DNA提取试剂盒(OMEGA D2485-01)提取植物基因组DNA。利用Nanodrop ND-2000微量核酸蛋白检测仪检测DNA质量及浓度。利用0.8%琼脂糖凝胶电泳检测DNA链的完整性,电泳缓冲液为 0.5×TBE。
1.2.2 ISSR引物筛选与PCR扩增 试验所选100条引物(UBC801~UBC900)由哥伦比亚大学公布,昆明硕擎生物科技有限公司合成。PCR反应体系为25 μL,包括模板DNA,引物2 μL,12.5 μL PCR反应混合物(Master Mix),加入双蒸水使体系达到25 μL。反应程序:94 ℃预变性3 min;94 ℃变性90 s, 50 ℃退火30 s, 72 ℃延伸1 min,43个循环;72 ℃延伸
从上述100条引物中初步筛选出28条进行退火温度梯度的试验,筛选出条带数量丰富、清晰稳定的引物12条,并确定最适退火温度。最后,利用这12条引物对16个铁线莲品种进行ISSR-PCR扩增,使用凝胶成像系统进行拍照分析。
1.2.3 数据记录与分析 根据凝胶成像结果,同一引物扩增出来的条带,在同一迁移率上出现条带记为“1”,丢失条带记为“0”,建立0、1二元数据矩阵。选择多态性位点比较丰富的引物,构建16个铁线莲品种的指纹图谱;利用NTSys2.1数据分析软件计算品种间的遗传距离和遗传相似系数,并采用UPGMA法构建其遗传聚类图。
2 结果与分析
2.1 铁线莲DNA提取效果
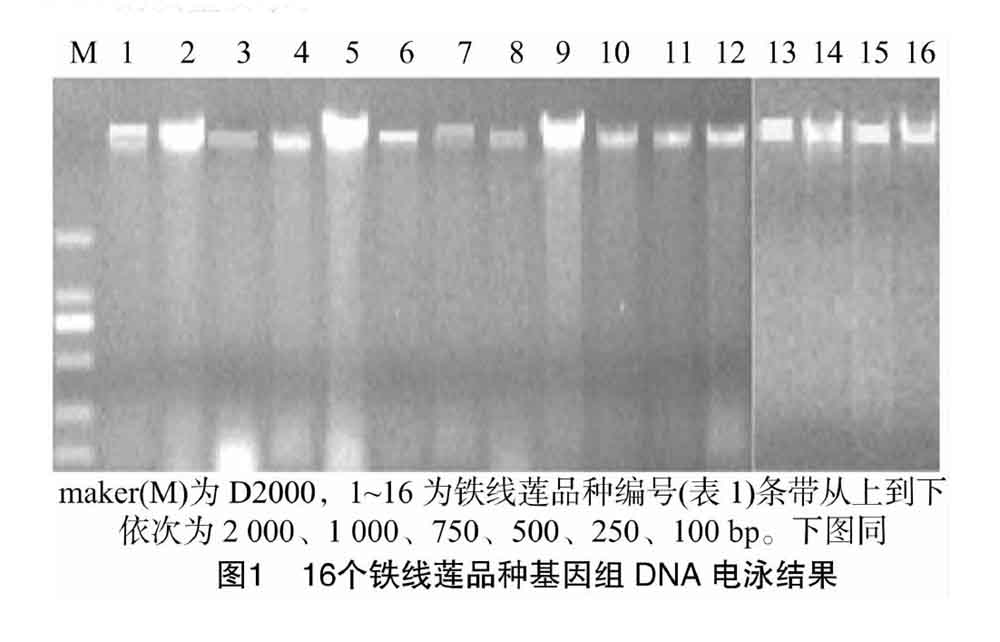
利用DNA提取试剂盒提取铁线莲16个品种的DNA都是一条完整清晰的谱带(图1),没有RNA污染、拖尾和弥散现象,利用超微量核酸蛋白测定仪测得DNA的浓度为55~120 ng/μL。提取的DNA可以满足ISSR-PCR反应对模板DNA的质量要求。
2.2 ISSR引物筛选及扩增结果
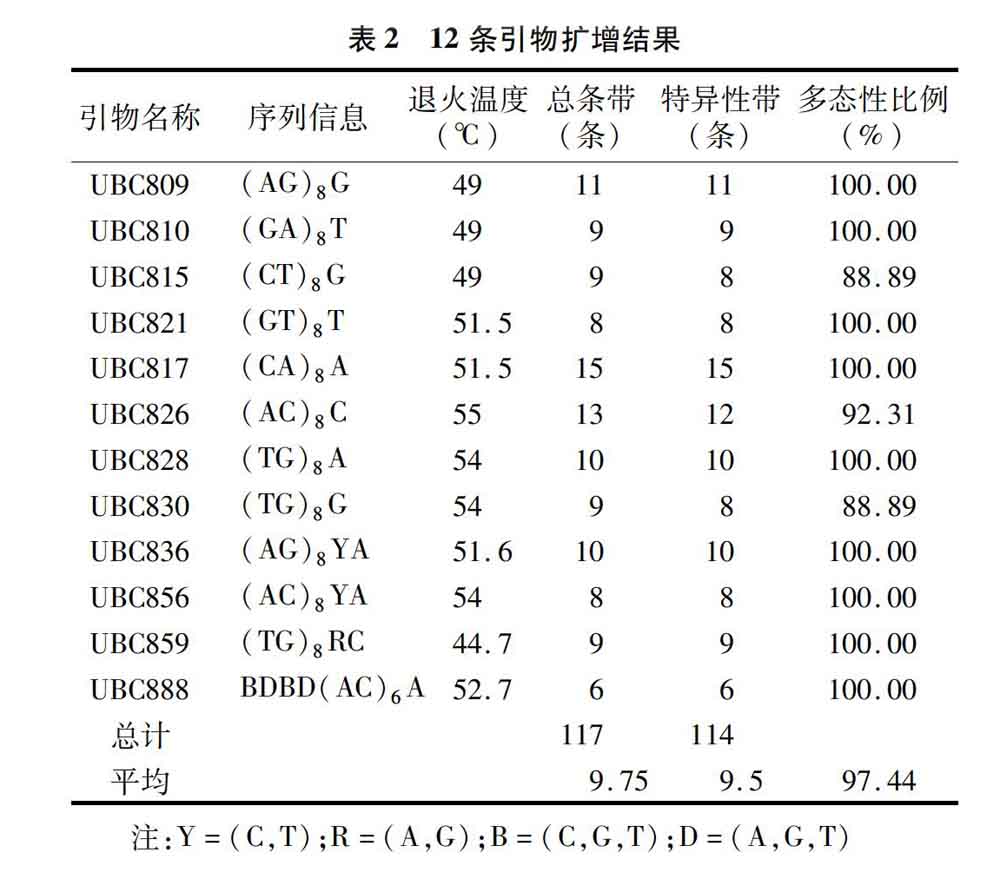
从100条ISSR引物中筛选出12条扩增条带清晰稳定、数量丰富的引物,分别为UBC809、UBC810、UBC815、UBC821、UBC817、UBC826、UBC828、UBC830、UBC836、UBC856、UBC859、UBC888。通过退火温度梯度(44.7~62.5 ℃)试验,确定了每条引物的最佳退火温度。利用筛选出的引物对16个供试材料进行ISSR-PCR扩增。12条引物共扩增出117条带,其中特异性条带共有114条,每个引物平均扩增出来9.75个位点,平均特异性位点达到9.5个,多态性比率达到97.44%(表2);扩增出的条带片段大小为250~1 500 bp。引物UBC817、UBC815和UBC856扩增结果见图2、图3、图4。
x2.3 16个铁线莲园艺品种的ISSR聚类结果
利用12条引物扩增出来的117谱带建立16个铁线莲品
种的遗传相似系数矩阵(表3),各品种之间的相似系数在 0.20~0.73之间,平均相似系数为0.51。说明铁线莲品种具有较高的遗传多样性,各品种间的遗传变异较大,环境适应能力较强。其中,包查德女伯爵和红衣主教相似系数最大,为0.73,这表明2个品种之间的亲缘关系相对较近;塞西尔和迈克莱特相似系數最小,为0.20,表明这2个品种的亲缘关系相对较远。从遗传相似系数来看,塞西尔同其他品种之间的遗传相似系数较小,为0.20~0.35,表明该品种与其他品种相比遗传变异大、亲缘关系远。
采用UPGMA法对供试的16个铁线莲园艺品种进行聚类分析,结果表明,在相似系数为0.50处,可将16份材料聚为3类(图5)。其中,塞西尔划为A类,紫铃铛划为B类,C类包括其余14个品种。在相似系数为0.53处,可将16份材料划分为4个类群。塞西尔划为第1类;紫铃铛划为第2类;第3类包括月光、迈克莱特、格拉芙泰美女和普鲁吐斯;其余10个品种划为第4类。
2.4 16个铁线莲园艺品种的ISSR指纹图谱构建
利用扩增出条带清晰、条带数量多、多态性丰富的引物构建16个铁线莲品种的遗传指纹图谱。利用引物UBC815和UBC817构建16个铁线莲品种的指纹图谱(表4),该指纹图谱可以有效地对这些铁线莲品种进行区分、鉴定。
3 结论与讨论
本研究利用ISSR分子标记技术筛选出的12条引物扩增出117个位点,多态性条带的比例为97.44%,每条引物扩增出的条带数为6~15条,多态性条带所占比例为88.89%~100%。其中12条引物中有9条引物扩增出的条带多态性达到了100%,说明铁线莲栽培品种具有较高的遗传多样性和复杂的遗传背景。利用引物UBC815和UBC817建立起的16个铁线莲品种的DNA指纹图谱可以对不同品种进行有效地区分和快速鉴定。对材料进行ISSR聚类分析,在相似系数0.50处可将16份材料聚为3类。A类只有品种塞西尔;B类只有紫铃铛;C类有普鲁吐斯、格拉芙泰美女、迈克莱特、月光、美佐世、天真一瞥、小鸭、钻石球、艾米莉亚花盘、蓝天使、包查德女伯爵、红衣主教、面白和吉恩斯。其中,包查德女伯爵与红衣主教之间的相似系数最大,为0.73,亲缘关系最近,遗传背景相似。从花朵形态特征来看,包查德女伯爵和红衣主教除花朵颜色不同外,都具有相同数量苞片,花期都为6—9月,花型都是平盘型,萼片中部具有竖状条纹,说明ISSR分子聚类与形态聚类在一定程度上的一致性。C类中铁线莲的花型都为平盘型,A类花型为长瓣型,B类为铃铛型。以此来看,按照花型分类也可以将材料划分为3类,这与ISSR聚类结果保持一致,拥有相同花型的品种之间可能具有相对较近的亲缘关系。但是在同一花型中,不同品种的花朵颜色、单重瓣及萼片数量等性状表现不一,这可能是铁线莲品种之间相互杂交导致种质资源混杂的结果,说明分子聚类结果与形态学分类结果存在一定的差异。塞西尔和迈克莱特之间的相似系数最小,为0.20,说明这2个品种之间的亲缘关系较远。在铁线莲新品种培育中,可以利用这2个品种进行杂交,可能会产生较好的杂种优势。
按照栽培品种的生境特点、开花特性及开花时间等,园艺家将铁线莲品种划为不同的品种类群。按照ISSR分子标记聚类结果将16份材料划分的3大类群中,A类只有1个长瓣型品种;B类只有1个单叶型品种;C类中包括8个早花大花型品种、1个德克萨斯型品种、2个意大利型品种和3个晚花大花型品种。ISSR聚类结果显示,不同园艺类型的铁线莲品种划分到同一类中,表明按照园艺性状分类结果与分子标记聚类结果存在一定的差异。这主要是因为园艺家对铁线莲品种的分类是为了园艺种植的考量,分类依据主要是植物生长环境、适应能力、花朵大小、颜色和开放时间等园艺特性。分子聚类与园艺分类的差异体现了铁线莲品种之间的遗传变异程度之大,这是长期人工选择和自然进化的共同作用的结果。ISSR聚类结果可以为铁线莲品种亲缘关系鉴定和杂交育种中亲本选择提供参考。与形态学标记相比,以遗传物质DNA为基础的ISSR分子标记重复性好,稳定性高,受环境因素影响小,在植物花卉种源信息鉴定、品种分类和遗传多样性评价等[10-12]方面具有广泛的应用。
参考文献:
[1]中国科学院中国植物志编委会.中国植物志(第28卷)[M]. 北京:科学出版社,2000.
[2]李同水. 藤本皇后铁线莲[M]. 长春:吉林科学技术出版社,2012.
[3]Gardner N,Hokanson S C. Intersimple sequence repeat fingerprinting and genetic variation in a collection of Clematis cultivars in commercial germplasm[J]. HortScience,2005,40(7):1982-1987.
[4]王 磊,王红梅,贾 君,等. 铁线莲属植物品种及野生种的物候期观察分析[J]. 浙江农业科学,2016,57(2):198-201.
[5]孟 雪,郝笑微,王 锦. 铁线莲8个小花品种的适生性比较[J]. 安徽农业科学,2011,39(35):21597-21599.
[6]高露璐,李林芳,马育珠,等. 铁线莲品种群的花期观赏性状分析[J]. 园艺学报,2017,44(5):921-932.
[7]任佳伦. 铁线莲品种开花生物学特性及核型分析[D]. 杭州:浙江农林大学,2016.
[8]孙正海,王 锦,李世峰,等. 滑叶藤ISSR-PCR反应体系建立及优化[J]. 云南农业大学学报(自然科学版),2012,27(5):746-750.
[9]王 楠,王 锦,李宗艳,等. 铁线莲园艺品种ISSR-PCR反应体系优化与引物筛选[J]. 北方园艺,2016(1):80-83.
[10]张俊杰,马 旭,廖 源,等. 昆明56个桂花品种ISSR分析[J]. 西部林业科学,2015,44(3):45-52.
[11]罗 清,卢业飞,於艳萍,等. 24份杜鹃属植物的ISSR分子鉴定[J]. 西北林学院学报,2016,31(3):154-158.
[12]苗美美,赵明明,刘绪明,等. 百合(Lilium spp.)杂交后代ISSR分析亲缘关系[J]. 分子植物育种,2016(2):437-441.王连喜,王 田,李 琪,等. 基于作物水分亏缺指数的河南省冬小麦干旱时空特征分析[J]. 江苏农业科学,2019,47(12):83-88.
- 网络订餐管理系统应用
- 计算机组装与维护中的常见问题及解决方法
- Oracle数据库技术在网络视听节目管理中的应用
- 新媒体传播特征背景下网络短视频广告发展契机研究
- 水面漂浮污染物突发事件应急处理系统设计
- 基于CMS内容管理和Web防篡改技术的门户网站系统
- 机动通信网络快速响应能力探讨
- 广播与网络视频直播融合问题探讨
- 一种基于大数据分析的广电网络多层次安全防护方案
- 微视频广告在网络传播中的策略分析
- 移动信息服务在高校图书馆中的应用研究
- 倒立摆系统的建模与控制研究
- 微信公众号对民生新闻的议程设置
- 南昌绳金塔数字虚拟还原的方式与价值研究
- 计算机软件数据接口的应用分析
- 邻近双丁字路口智能互联交通灯控制系统
- 自媒体传播对武警部队执勤工作的主要影响及应对策略
- 广播电视工程中计算机技术的应用
- 大数据分析技术及其在医疗领域的应用
- 群体智能算法在机器学习当中的应用
- 机器视觉检测与人眼检测对比研究
- 2015年以来网络综艺节目现状与发展趋势浅析
- 网络视频广告效果的提升策略研究
- 网络社会负面事件对大学生价值观的影响及教育对策
- 基于单片机啼哭频率采集的分析
- overoffensive
- overoffensively
- overoffensiveness
- overoffensivenesses
- overoffice
- overofficious
- over-officious
- overofficiously
- overofficiousness
- overofficiousnesses
- overold
- overoperate
- overoperated
- overoperates
- overoperating
- over-opinionated
- overopinionated
- overopinionatedly
- overopinionatedness
- overopinionatednesses
- over-optimism
- overoptimisms
- overoptimist
- over-optimistic
- overoptimistic
- 令人喜爱
- 令人喷饭
- 令人失望不满的事
- 令人害怕让人害怕
- 令人心惊的浪涛
- 令人心惊胆颤
- 令人忧愁苦闷的局面
- 令人快慰
- 令人怜悯
- 令人悔恨的事
- 令人悲痛
- 令人惊叹的精美绝伦之物
- 令人惊叹绝倒
- 令人惊异
- 令人惊讶地突现
- 令人惊讶的事突现
- 令人惊骇的诗人
- 令人惋惜
- 令人感念
- 令人愤怒到极点
- 令人捧腹
- 令人掩鼻
- 令人敬畏的样子
- 令人敬畏的气派和威严完全丧失
- 令人极悲痛