李金霞 储博彦 赵玉芬 尹新彦 赵振峰

摘要:以8个白桦家系为试验材料,在大田栽植的条件下,测定其在5—9月叶片的相对电导率、MDA含量、SOD活性和可溶性蛋白含量共4个生理指标,并利用隶属函数法对其越夏期的耐热性进行综合评价。结果表明,4个生理指标均表现出与气温变化的正相关,8个家系均能在石家庄地区安全越夏,但耐热性有差异,其中家系CD和1-41耐热性最强,家系1-59和1-46的耐热性最差。
关键词:白桦;家系;耐热性;综合评价
中图分类号: S792.153.01;Q945.78 ?文献标志码: A ?文章编号:1002-1302(2020)17-0148-05
温度是影响植物生长发育的重要因子。在植物引种过程中,从高纬度向低纬度、高海拔向低海拔引种,夏季高温成为引种成功与否的重要限制因子,而在城市绿化中,由于城市热岛效应的存在以及全球气候变暖的加剧,城市园林绿化也迫切需要新的耐热型植物品种。因此研究植物在越夏期高温下的生理生化反应,以耐热性为评价指标对引种植物的抗逆性进行评价,有助于筛选出优良的耐热型植物品种,对丰富城市园林景观也具有重要意义。
白桦(Betula platyphylla Suk.)为桦木科桦木属落叶乔木,其枝叶轻盈,秋叶金黄,树皮洁白雅致,在园林景观配置中具有独特的观赏效果。耐热型白桦属于高海拔树种,喜冷凉湿润气候。关于白桦的抗逆性生理已有部分研究,但多集中在抗干旱[1]和抗低溫胁迫[2-3]方面。有学者对耐热型白桦的种子筛选[4]、栽培基质[5]和育苗[6]、栽培技术[7]进行了研究,并在人工培养箱内对白桦幼苗进行高温胁迫试验[8-9]。然而植物的耐热性是其在夏季自然条件下经受多种复杂因素胁迫的综合表现[10],目前对大田栽植条件下的白桦幼苗在越夏期的生理表现尚未见报道。为丰富石家庄城市绿化的树种选择,2015年从东北林业大学和承德共引进耐热型白桦家系8个,并于2017年进行田间耐热性测定,以期筛选出最具耐热潜力的优良家系,从而为白桦在石家庄的引种奠定基础。
1 材料与方法
1.1 试验材料
2015年从东北林业大学引进耐热型白桦优良家系7个(1-39、1-41、1-46、1-54、1-59、1-56、2-5),从承德引进耐热性白桦家系1个(CD),共计8个家系。2015年春季进行育苗,2016年春季分栽于河北省林业科学研究院试验基地,试验期间进行相同的常规栽培管理。
1.2 试验地点
试验地点位于石家庄市河北省林业科学研究院试验基地(114°28′34.03″E、38°8′25.67″N),试验期间气温情况见表1。
1.3 试验方法
2017年5月在8个白桦家系中各选出3株高度与基径接近平均值的标准株,分别于5月13日、6月16日、7月11日、8月25日和9月21日的14:00取标准株枝条中上部成熟叶片,对其进行田间耐热性测定,所有测定均重复3次。测定当天及前2 d均为晴天,具体气温见表1。测定指标包括相对电导率、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、可溶性蛋白含量,其中MDA含量采用硫代巴比妥酸法测定,SOD活性采用氮蓝四唑(NBT)光还原法测定,可溶性蛋白含量采用考马斯亮蓝G-250染色法测定。
1.4 耐热性评价
采用隶属函数法对不同白桦家系的耐热性进行综合评价。与耐热性呈正相关的参数采用式(1)进行计算,与耐热性呈负相关的参数采用式(2)进行计算。
式中:U(Xijk)为第i个家系第j个月份第k项指标的隶属度,且U(Xijk)∈[0,1],Xijk为第i个家系第j个月份第k项指标的测定值,Xkmin和Xkmax分别为所有家系中第k项指标的最小值和最大值。用每个家系各项指标隶属度的平均值作为评价其耐热性的依据。
2 结果与分析
2.1 不同白桦家系相对电导率的月变化
相对电导率能够反应逆境对植物组织的伤害程度,是比较理想的植物抗逆性鉴定的生理指标之一[10]。由表2可以看出,5—9月不同白桦家系相对电导率均表现为先升高后下降,与气温表现出一致的变化规律。5月,不同白桦家系的相对电导率在9.12%~17.20%。7月,所有白桦家系电导率均达到最大值,与5月相比,家系1-59的增长率最大,为155.68%,同时也是7月电导率值最大的家系,说明该家系膜结构的受损程度最大。8、9月,当气温下降后,所有家系相对电导率也随之降低。
2.2 不同白桦家系MDA含量的月变化
MDA含量是植物是否氧化过度的判定标准,MDA含量越高,说明质膜受损越严重。5—9月不同白桦家系MDA含量表现为先升高后下降,除家系2-5和1-59在8月MDA含量最高外,其余家系均在7月达到最大值。5月,不同白桦家系的MDA含量在4.06~16.50 nmol/g,其中家系1-56和CD的MDA含量明显低于其他家系,但在7月的增长率却明显高于其他家系,分别达到5月的5.39倍和5.56倍,说明气温的升高对这2个白桦家系MDA含量的积累影响更为明显。9月气温下降后,MDA含量也随之降低,但除家系 1-39外,其他家系均不能恢复到5月份的水平,尤其是家系1-56和CD(表3)。
2.3 不同白桦家系SOD活性的月变化
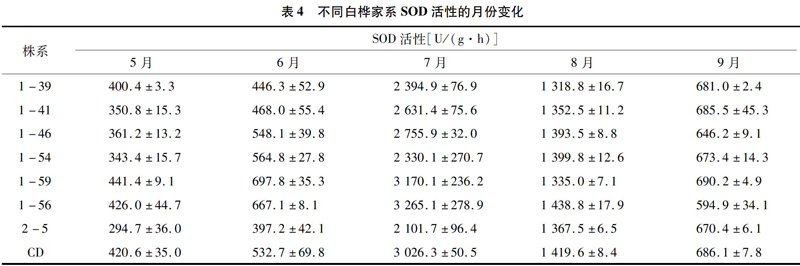
SOD是植物体内清除活性氧自由基的关键酶,在保护细胞免受氧化损伤过程中具有十分重要的作用。5月,不同白桦家系的SOD活性在 294.7~441.4 U/(g·h),6月气温为34.8 ℃时,与5月增长幅度不大,但在7月气温达到 38.8 ℃ 时,所有家系的SOD活性均成倍数增加,达到2 101.7~3 265.1 U/(g·h),8、9月气温下降,高温胁迫得到缓解,SOD活性也随之降低,但仍维持在较高的水平(表4)。
2.4 不同白桦家系可溶性蛋白含量的月变化
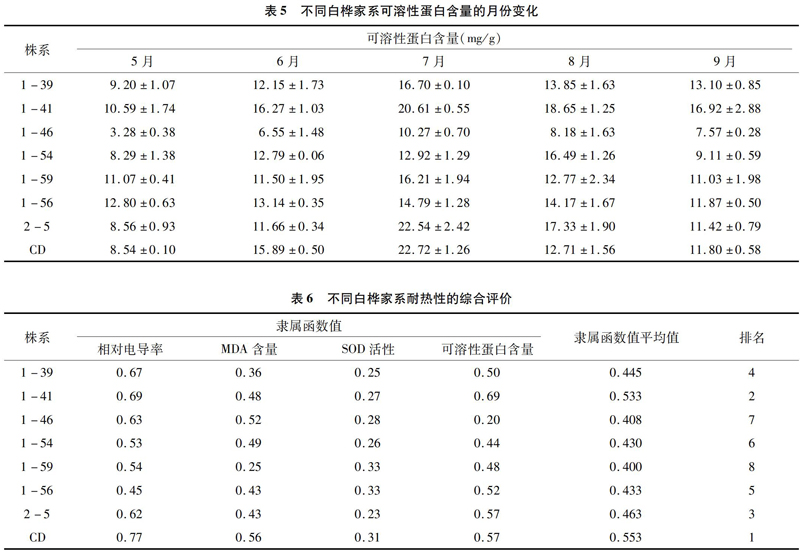
可溶性蛋白是植物细胞内常见而有效的渗透调节物质,耐热植物在高温胁迫下可保持较高的蛋白质合成速率和较低的蛋白质降解速率[11],从而维持较高的可溶性蛋白含量。从表5可以看出,不同白桦家系的可溶性蛋白含量差异较大,5—9月家系1-41可溶性蛋白含量平均值最高,为16.61 mg/g,家系 1-46 最低,为7.17 mg/g。除家系1-54外,其余均在7月达到最大值,表现出与气温变化的正相关。
2.5 不同白桦家系耐热型的综合评价
采用隶属函数法,对每个家系各指标的隶属函数值求平均值,以此为据对其耐热性进行评价。对与耐热性呈负相关的相对电导率和MDA含量采用公式(2)进行计算,对与耐热性呈正相关的SOD活性和可溶性蛋白含量采用公式(1)进行计算,计算结果见表6。从表6评价结果可以看出,家系CD和1-41的耐热性最强,而家系1-59和1-46的耐热性最差。
3 结论与讨论
细胞膜是植物细胞与外界环境的屏障,维持着细胞内环境的稳定性,当植物处于逆境时,细胞膜受到损伤往往会导致膜透性增大,电解质从细胞内渗透出来,引起组织浸出液电解质浓度随之增高,相对电导率升高。因此,相对电导率反应了细胞膜的稳定性,被广泛应用于多种植物的耐热性鉴定,电导率越大,说明细胞膜受到的伤害越大,其耐热性就越弱[9,11-14]。在本试验中,5—7月,不同白樺家系的电导率均随着气温的升高而增加,然而不同家系的变化幅度不同,说明家系间的耐热性差异在细胞膜透性的增加幅度上表现较为明显。7月过后,气温下降,电导率也随之降低,说明在石家庄地区,7月高温并没有使白桦各家系的细胞膜发生不可逆的损伤。
MDA是膜脂过氧化作用的产物,其含量可以表示膜脂过氧化的程度和植物对逆境条件反映的强弱。许多研究表明,MDA含量与植物的耐热性呈现出负相关,在高温胁迫下,耐热性强的植物,仍能维持较高的保护酶活性,清除活性氧的能力较强,MDA积累较少;而耐热性弱的植物,当受到重度的高温胁迫时,则易造成大量的MDA等有害物质的积累[11-16]。本研究结果与其他学者研究较为一致,5—7月随着气温升高,6个白桦家系的MDA含量逐渐积累,在7月气温最高时达到最大值。之后随气温下降,MDA含量逐渐降低。然而家系1-59和2-5的MDA含量却在8月达到最大值,9月有所降低,这一现象可能是受到复杂的大田环境的影响,原因有待进一步研究。
高温胁迫引起的膜脂过氧化过程中,氧自由基O-2·、H2O2等有毒物质的产生速度与保护酶系统在高温下的活性共同决定着植物的耐热性[17],而SOD是植物体内清除活性氧自由基的关键酶,它可以减轻膜脂过氧化程度,保持膜系统的稳定性。本研究中,随着气温升高,6月所有家系SOD活性增长幅度较小,然而到了7月,均出现呈倍数增加,说明白桦在高温胁迫下,会通过增强SOD活性,来减轻植物体内活性氧平衡的破坏,提高对高温的适应性。这与胡佳佳等对屋顶绿化植物的耐热性评价结果[18]相一致。
作为植物细胞内重要的渗透调节物质,可溶性蛋白对细胞膜的结构与功能稳定具有重要的作用[19]。研究表明,高温导致氮素代谢失调是植物生长受到影响的一个重要原因[11]。耐热性较强的植物在高温条件下合成蛋白质的速率较高,降解速率较低。本研究中,虽然不同白桦家系可溶性蛋白含量变化均表现出与气温的正相关性,夏季高温使白桦各家系保持较高的蛋白质合成速率和较低的降解速率,气温降低后,又基本回归到正常状态。但不同家系间的含量差异较大,且在不同月份的变化幅度也表现出差异性。
目前大多学者对植物耐热性的研究多是在人工气候培养箱中模拟高温胁迫,这种各因素可控的试验条件难以准确反映出自然界的复杂环境对植物生长的影响,同时由于植物不同品种之间潜在的耐热性差异需经过一定的热锻炼才能充分表现出来[20],因此不能够准确地反映植物的耐热性。本研究通过大田试验,在自然环境下进行,8个白桦家系经过顺应气候的热锻炼,能够真实表现出各自耐热性的强弱。目前在植物耐热性研究中,温度梯度的设置多为单调升高,但是对高温胁迫后温度降低时植物的生理反应则报道较少,本研究当中,8、9月气温降低,各个家系的4个生理指标均有不同程度的恢复,说明在石家庄地区,7月高温并未对白桦的生长造成不可逆的伤害。
耐热性是植物在夏季经受多种因素胁迫下的一种综合表现,是一个综合性指标,而且不同植物家系品种在耐热机制上也存在差异[21],因此使用单一指标并不能全面地反应植物的耐热性强弱,而应该使用多个指标进行综合评价。本研究通过隶属函数法,以相对电导率、MDA含量、SOD活性和可溶性蛋白含量为评价指标,综合评价了8个白桦家系的耐热性。结果表明,8个白桦家系均能在石家庄安全越夏,其中家系CD和1-41耐热性最强,家系1-59和1-46的耐热性最差。
参考文献:
[1]孙国荣,彭永臻,阎秀峰,等. 干旱胁迫对白桦实生苗保护酶活性及脂质过氧化作用的影响[J]. 林业科学,2003,39(1):165-167.
[2]杨德浩,杨敏生,王进茂,等. 欧洲白桦苗期低温胁迫时膜系统的变化[J]. 东北林业大学学报,2004,32(6):13-15.
[3]乌凤章,王柏臣,刘桂丰,等. 低温胁迫对白桦幼苗生长和生理的影响[J]. 东北林业大学学报,2008,36(9):8-10.
[4]卜燕华,王永格,李子敬,等. 耐热性白桦种子筛选研究[J]. 北京园林,2014,30(3):30-33.
[5]王永格,卜燕华,李子敬,等. 不同栽培基质对耐热白桦生长的影响[J]. 北京农学院学报,2014,29(3):73-77.
[6]卜燕华,王永格,李子敬,等. 北京地区不同种源白桦育苗试验初报[J]. 北京农学院学报,2015,30(1):81-85.
[7]任春生,卜燕华,王永格. 耐热白桦栽培技术[J]. 北方园艺,2016(24):76-79.
[8]郝 征,张 钢,李 亚. 不同高温胁迫对白桦幼苗几个生理生化指标和电阻抗图谱参数的影响[J]. 西北植物学报,2010,30(9):1844-1851.
[9]李子敬,王永格,卜燕华,等. 北京园林绿化耐热白桦种源筛选[J]. 科学技术与工程,2013,13(16):4635-4638,4646.
[10]王 涛,田雪瑶,谢寅峰,等. 植物耐热性研究进展[J]. 云南农业大学学报,2013,28(5):719-726.
[11]郭思佳,冯永文,王 兵,等. 4种野生地被植物的耐热性试验[J]. 西部林业科学,2015,44(4):96-99,104.
[12]欧祖兰,曹福亮,郑 军. 高温胁迫下银杏形态及生理生化指标的变化[J]. 南京林业大学学报(自然科学版),2008,32(3):31-34.
[13]施 曼,祝遵凌. 越夏期不同欧洲鹅耳枥品种耐热性评价[J]. 西北林学院学报,2015,30(6):59-64.
[14]胡伟娟,张启翔,潘会堂,等. 报春花属植物耐热性生理指标研究初探[J]. 中国农学通报,2010,26(5):158-163.
[15]张文娟,李连国,郭金丽,等. 高温胁迫下6种景天植物的形态特征及生理响应[J]. 内蒙古农业大学学报(自然科学版),2012,33(4):41-45.
[16]李长林,杨守坤,金 莉,等. 高温胁迫对北美海棠金吉丽和特丽的生理指标影响[J]. 中国农学通报,2014,30(10):170-173.
[17]刘祖祺,张石城. 植物抗性生理学[M]. 北京:中国农业出版社,1994.
[18]胡佳佳,裴宗平,刘 田,等. 几种常用屋顶绿化植物的耐热性评价[J]. 环境科学与技术,2011,34(1):187-190,205.
[19]袁启凤,李仕品,严佳文,等. 不同贮藏温度对火龙果紫红龙可溶性蛋白质含量和抗氧化酶活性的影响[J]. 江苏农业科学,2019,47(2):194-197.
[20]Paulsen G. Application of physiology in wheat breeding[J]. Crop Science,2002,42(6):2228.
[21]張朝阳,许桂芳. 利用隶属函数法对4种地被植物的耐热性综合评价[J]. 草业科学,2009,26(2):57-60.
- 将项目式学习引入小学数学课堂的实践与思考
- 班级学习小组建设和小组评价策略研究
- 巧绘思维导图 提高阅读质量
- 浅析初中英语有效阅读教学模式的构建与运用策略
- 深化文本解读 优化高中英语阅读课堂教学
- 浅析英语配音软件在初中英语教学中的应用策略
- 思维导图在高中英语写作教学中的应用策略探究
- 激活初中英语阅读高效课堂的方法探究
- 高中文言文单元群文阅读教学模式设计初探
- 小学语文教学和班主任管理的有效结合思考
- 关于提高小学语文阅读教学有效性策略的探究
- 生命关怀教育在小学语文教学中的应用策略探述
- 浅谈如何在高中语文教学中渗透中华优秀传统文化
- 初中语文阅读教学与学生阅读能力的培养策略分析
- 浅析小学语文个性化学习中信息技术的整合实践运用
- 情境教学法在高年级小学语文教学中的作用分析
- 小学语文写作教学分段训练模式分析
- 用创新理念点亮小学语文课堂教学的策略分析
- 探究情景教学法在语文课堂教学中的应用策略分析
- 前置性学习模式在农村小学语文课堂的运用策略探索
- 课堂辩论在高中语文教学中的应用策略探究
- 基于新时代背景下以生活热点为切入口的“育人型微写作”指导
- 关于提高初中语文课堂教学有效性的策略探究
- 小学语文教学中海量阅读与阅读素养的提升策略分析
- 浅谈如何提高初中语文阅读教学的有效性
- elucidated
- elucidates
- elucidating
- elucidation
- elucidations
- elucidative
- elucidator
- elucidators
- elucidatory
- elude
- eluded
- eluder
- eluders
- eludes
- eludible
- eluding
- elusive
- elusively
- elusiveness
- elusivenesses
- elven
- elves
- 'em
- emaciated
- emaciation
- 群臣百官
- 群花
- 群花生树
- 群芳
- 群芳斗艳
- 群芳竞艳
- 群芳髓
- 群英
- 群英会
- 群英荟萃
- 群莺竞飞
- 群萃
- 群萌
- 群落
- 群虫飞动声
- 群虱处裈中
- 群蚁溃堤
- 群蚁能降毒蛇
- 群蚁趋膻
- 群蚁附膻
- 群蛀
- 群蛙叫声
- 群蛙鼓吹
- 群蜂簇拥
- 群行