刘君洋 王明力 赵来朋282
摘要:综合分析艾比湖流域胡杨冠下土壤有机碳(SOC)、全氮(TN)、全磷(TP)分布及生态化学计量特征,对土壤SOC、TN、TP含量及C/N、N/P、C/P的水平和垂直变化进行单因素方差分析,用最小显著性差(LSD)对数据进行多重比较。结果表明,从肥岛外至冠幅中央,随着距离的增加肥岛效应不断增强,SOC、TN含量显著增加(P<0.05),0~5、5~10、10~15 cm这3个土层深度下SOC和TN含量分别增加85%、83%、83%和87%、75%、78%,而TP含量变化不显著;随着深度的增加,TN、SOC和TP含量都表现出减小的趋势,0~5 cm的TN、SOC含量显著大于10~15 cm(P<005),各土层深度下TP差异均不显著;从肥岛外至冠幅中央随着肥岛效应不断增强,0~5、5~10、10~15 cm这3个土层的N/P和C/P显著增加(P<0.05),分别增加83%、66%、74%和80%、77%、80%,而C/N变化不明显;随着深度的增加,N/P和C/P都呈减小趋势,0~5 cm的N/P显著大于10~15 cm(P<0.05)。说明胡杨冠下土壤C、N变化显著,空间异质性强,表现出明显的肥岛效应,但P变化不显著;N/P和C/P受肥岛效应显著影响,随着肥岛效应的增强明显增加,而C/P几乎不受影响;胡杨冠下肥岛的P素有效性较高,有机质矿化分解速率较快,同时表现出较为严重的N素限制。
关键词:生态化学计量;肥岛;胡杨;艾比湖流域
中图分类号: S718.5? 文献标志码: A? 文章编号:1002-1302(2020)20-0282-06
土壤理化性质通常存在空间异质性,特别是在干旱区生态系统中,突出表现为植物下的肥岛效应[1]。肥岛效应是土壤养分资源在植物冠幅下的聚集,会对植物-土壤之间的碳(C)、氮(N)、磷(P)循环与转化产生重要影响[2],而C、N、P是生命体三大元素,这些元素存在相对稳定的比率,其动态变化决定了生命体和生态系统的特征[3],对揭示养分的有效性及平衡机制具有重要的科学意义[4]。因此,立足于干旱区植物的肥岛效应对生态化学计量展开研究,有助于了解植物养分的可获得性,评估植被健康状况,促进干旱区生态保护。
生态化学计量学在研究生态系统养分循环方面备受重视[5],而外部因素对生态化学计量的影响也受到越来越多的关注[6-7]。Wei等发现,长期植树造林会明显提高土壤中的有机碳、全氮、全磷含量,C/P、N/P会随植树造林时间的增加而增加[8]。高丽倩等探究了生物结皮对生态化学计量的影响,发现土壤化学计量特征与生物结皮生物量显著相关,C/N随演替变化较小,而C/P和N/P随结皮演替明显增加[9]。Xiao等发现,随着海拔升高,C/P和N/P表现出先增加后减少的趋势,生态系统中的限制性养分也受到海拔梯度的影响[10]。此外,还有学者研究了纬度[11]、土壤类型[12]等对生态化学计量的影响,并取得了一定的成果。然而,作为一个重要的物理和生物學因素,肥岛效应如何影响土壤生态化学计量却鲜有报道。因此,探讨肥岛效应影响下的生态化学计量特征变化,有助于阐明生态化学计量学的影响因素和机理,对研究干旱荒漠生态系统土壤的C、N、P生物地球化学循环具有重要意义。
近50年来,艾比湖流域生态环境急剧恶化,已成为北疆尘暴重要的策源地,土壤沙化盐化加剧[13]。胡杨(Populus euphratica Oliv.)作为艾比湖流域主要建群种之一,同时也是新疆荒漠生态系统中乔木的典型代表[14],对保持艾比湖流域乃至荒漠区生态系统的平衡起着重要作用。基于此,本研究以胡杨为研究对象,探讨肥岛的特征及其对生态化学计量的影响。这将有助于对C、N、P生物地球化学循环的解释[15],对丰富干旱区植物冠下土壤的肥岛效应和生态化学计量理论具有重要意义。
1 研究区域与研究方法
1.1 研究区概况
艾比湖流域地处欧亚大陆腹地,准噶尔盆地南隅,地理位置79°53′~85°02′E、43°38′~45°52′N,流域面积50 621 km2。该区域属典型温带大陆性干旱气候,冬季寒冷漫长,夏季炎热难耐,植被稀疏,生态环境脆弱[16],年降水量100 mm左右,年蒸发量 1 600 mm 以上,日照时数约2 800 h,极端最高气温44 ℃,极端最低气温-33 ℃,年平均气温为6~8 ℃。植被类型除优势种胡杨(Populus euphratica Oliv.)、梭梭[Haloxylon ammodendron (C.A. Mey.) Bunge]外,还伴有柽柳(Tamarix ramosissima Ldb.)、芦苇[Phragmites australis (Cav.) Trin. ex Steud.]、盐豆木[Halimodendron halodendron (Pall.) Voss.]、沙拐枣(Calligonum ebinuricum Ivanova.)等。
1.2 样品采集
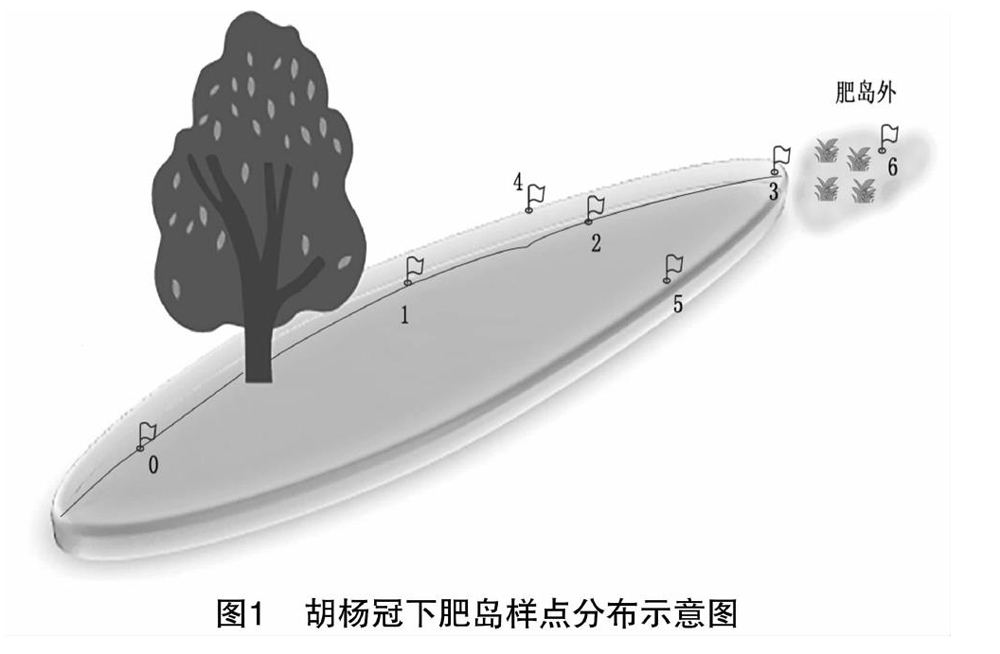
2017年在艾比湖流域进行土壤样品的采集,以胡杨冠下肥岛为研究对象,选择胡杨胸径、树高、冠幅等生长情况较为一致的5个样地,同时确保该样地距离最近的胡杨冠幅边缘至少5 m。对选取的每个样地,从冠幅中央至肥岛边缘水平线上取3个点,分别用1、2、3表示,在点2两侧另取2点与点3共同确定肥岛边缘养分值,用4、5表示,采集肥岛外土样,用6表示。此外,在上风向胡杨冠下裸斑面积较小一侧采集一个与点1在同一水平线上且与树干等距的土样,用0表示(图1)。在每个采样点以5 cm为间隔采取0~15 cm土样,共105(5×7×3)个。
1.3 样品测定及数据分析
土壤样品在室内自然风干、研磨和过筛处理后,进行土壤有机碳(SOC)、全氮(TN)和全磷(TP)的分析测定。有机碳含量采用重铬酸钾容量法-外加热法测定;全氮含量采用凯氏定氮法测定;全磷含量采用HClO4-H2SO4消煮钼锑抗比色法测定。采用Microsoft Excel 2007和SPSS 19.0进行数据处理和分析。采用方差分析(ANOVA)分析土壤C、N、P含量及C/N、N/P、C/P的水平和垂直差异,用最小显著性差(LSD)对数据进行多重比较。
2 结果与分析
2.1 胡杨冠下土壤有机碳(SOC)、全氮(TN)和全磷(TP)的含量变化
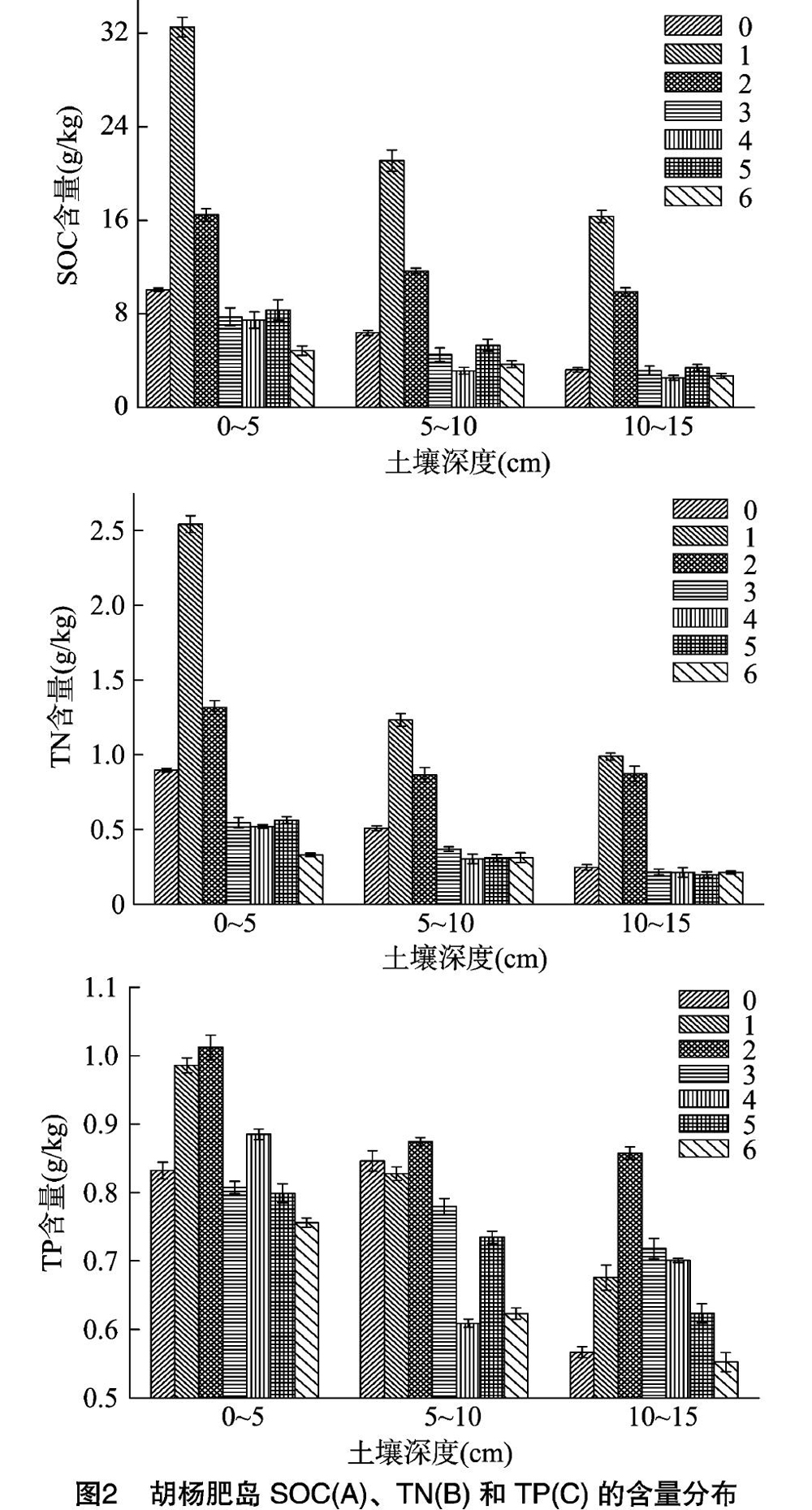
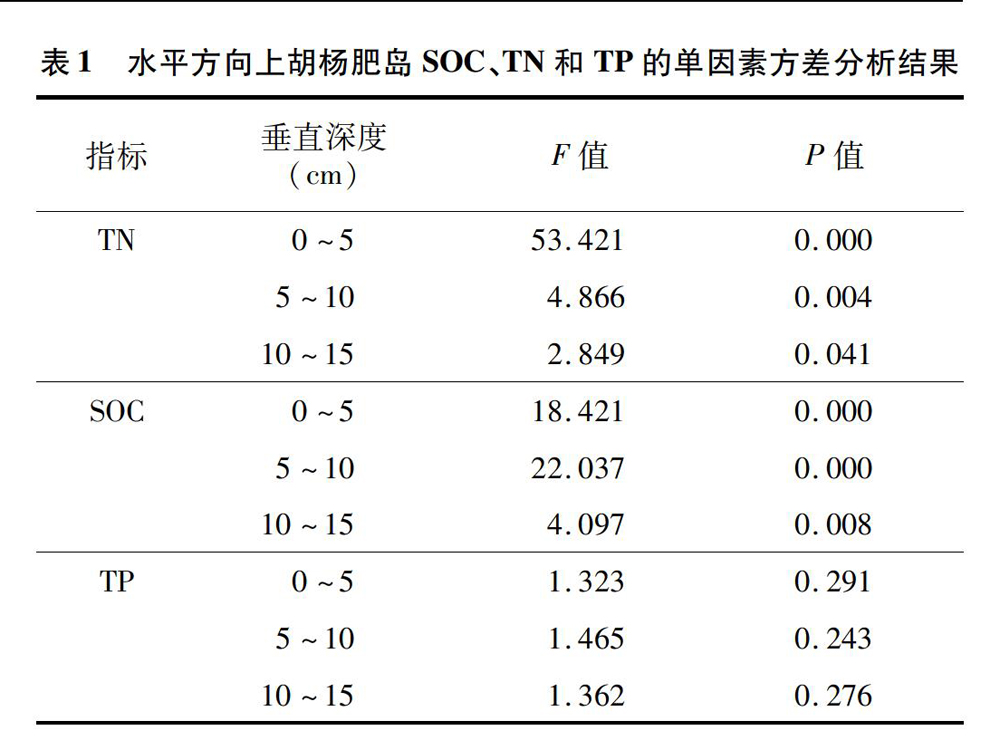
从图2-A、图2-B可以看出,全氮和有机碳的含量变化较为一致,在0~5、5~10、10~15 cm上都表现出相似的趋势。从冠幅中央至肥岛外,全氮和有机碳的含量都显著下降(表1),尤其从点1到冠幅边缘的点3,有机碳含量分别减少76.18%、7863%、80.62%,全氮含量则减少78.55%、7011%、7814%,但点3之后含量趋于稳定,点3与点4、点5之间无显著差异,说明肥岛边缘值是稳定的,至肥岛外的点6处,3个土层的全氮含量已分别降低至0.33、0.31、0.21 g/kg,有机碳含量則降低至8.39、6.36、4.66 g/kg。可以看出,全氮和有机碳的含量随着距离的变化表现为明显的下降趋势,呈现出典型的肥岛特征。从图2-C可以看出,与全氮和有机碳不同,全磷含量随着距离的变化呈现出先增加后减小的趋势,且并未产生明显的差异,因此,全磷含量的最高值并未像全氮和有机碳一样出现在点1,而在点2。基于此,全磷含量介于0.55~1.01 g/kg,受肥岛效应影响不显著,存在一定的稳定性。
由图2可见,在垂直方向上,从点1到点6采样点的全氮、有机碳和全磷含量最高值都在0~5 cm土层,且都呈现出了随深度增加而减小的趋势。由表1可见,全氮与有机碳的变化趋势相同,0~5 cm的显著大于10~15 cm的(P<0.05),5~10 cm和10~15 cm之间差异不显著。全磷含量在0~5、5~10、10~15 cm之间的差异都不显著。另外, 对点0与其他各点的养分进行方差分析发现,点0的全氮和有机碳含量在3个土层上均显著小于点1(P<005),而与肥岛外的点6无显著差异,说明冠幅两侧的养分差异很大,上风向胡杨冠下裸斑面积较小一侧肥岛发育程度比另一侧低。
2.2 胡杨冠下土壤C/N、N/P、C/P变化
由图3可见,N/P和C/P值的变化与全氮、全磷的变化十分相似,在3个土层都具有下降趋势。从点1至点3迅速下降,3个土层的N/P分别下降73.82%、68.27%、79.42%,C/P分别下降了7092%、77.32%、81.76%,但在点3之后变化较小,各点之间无显著差异。通过分析(表2)表明,N/P和C/P随着距离冠幅中央的距离增加,3个土层在水平方向上都表现出显著差异(P<0.05),其值随距离的变化不断减小,这说明在水平方向上胡杨冠下土壤N/P和C/P受到了肥岛效应的显著影响。不同于N/P和C/P,C/N值并没有随着与冠幅中央距离的增加表现出明显的变异。3个土层的C/N最大值都在点5出现,略高于点1,5~10、 10~15 cm层的C/N值随着距离的变化,分别在 10.33~17.16和11.31~17.16间呈现波动变化,而0~5 cm层则呈现出缓慢上升的趋势,到点5达到最大值14.801。这表明C/N具有一定的稳定性,几乎不受肥岛效应的影响。分析肥岛未发育区的点0与肥岛内的点1、肥岛外的点6发现,点0的 N/P 和C/P值与点1在0~5、5~10、10~15 cm这3个土层上都表现出明显的差异,但与肥岛外的点6却无显著差异,说明在水平方向上胡杨冠下裸斑面积较小一侧与另一侧的生态化学计量存在显著差异。
另外,由图3可以看出,从胡杨冠下肥岛内的点1到肥岛外的点6,C/P和N/P在垂直方向上受肥岛效应影响而随深度变化。C/P和N/P的最高值都在0~5 cm且随深度的增加逐渐减小,其中0~5 cm 的N/P显著大于10~15 cm(P<0.05)。与C/P和N/P不同,各点的C/N在垂直方向上没有表现出一致的规律,点2、点5随深度增加先增大后减小,点3、点4、点6则先减小后增大,只有点4和点6的最大C/N值在0~5 cm,其他各点的最大值分别在5~10、10~15 cm出现,这表明C/N在垂直方向上具有一定的稳定性,不受土层深度的影响。在点0处,C/N随深度的增加而增加,但3个土层之间差异不显著,而C/P和N/P随深度的增加而减小,其0~5 cm分别与5~10、10~15 cm存在显著差异。
2.3 胡杨肥岛有机碳、全氮、全磷与N/P、C/P、C/N相关性分析
由表3可见,除了全磷与C/N之间关系不显著外,有机碳、全氮分别与N/P、C/P存在较高的相关系数,同时,全氮与C/P、有机碳与N/P较为一致的变化趋势也证明了养分会对生态化学计量产生一定的影响,而这也可以说明从肥岛外至冠幅中央 N/P、C/P显著增加、C/N变化不明显的原因。
3 讨论与结论
干旱区植物的肥岛效应会导致养分在水平和垂直方向上产生明显的空间异质性[17]。根据克伦梅德森和巴斯以土壤全氮为主要养分指标建立的肥岛模型,养分含量呈现出从冠幅中心向外逐渐下降的趋势,而随着土壤深度的增加,这一趋势逐渐下降[18]。本研究得到了相同的结果,在干旱区荒漠生态系统中胡杨下的土壤养分,特别是C和N,不仅从冠幅中心到肥岛外空地表现出明显的空间变异,而且从表层土壤到下层土壤都呈现出下降的趋势。这种空间异质性说明胡杨或许对土壤C、N、P的循环,尤其是其中的限制性营养元素起着严格的控制作用[19],它为胡杨尽可能地利用这些土壤养分要素提供了帮助,提高了胡杨适应贫瘠生境的能力。胡杨冠下裸斑养分差异很大,下风向面积较大一侧的养分含量明显大于另一侧,这可能是由于长期风向和胡杨树冠结构等因素长时间影响造成的[20-21]。胡杨肥岛效应对土壤养分具有明显的富集作用,而养分的变化会对生态化学计量产生一定的影响。Aponte等的研究结果表明,C/P主要受到全氮的影响,N/P主要受到有机碳的影响,而C/N主要受到全磷的影响[22]。本研究得到了类似的结果,除了全磷与C/N之间关系不显著外,有机碳、全氮分别与N/P、C/P存在较高的相关系数。
本研究中,胡杨冠下肥岛的全氮、有机碳与全球、全国的平均水平存在差异。胡杨冠下的全氮含量为0.20~2.54 g/kg,大部分样点的氮含量低于全球水平(2.1 g/kg)[23]和中国水平(2.3 g/kg)[24],表明处于干旱区的胡杨生长环境存在一定程度的氮养分限制,而氮素的缺乏是限制植物生长的一项关键因素,会影响叶绿素的组成和形成,最终影响植物生长和新陈代谢进程[25]。有机碳与全氮相同,大部分样点值比全球水平(25.71 g/kg)和中国水平(2951 g/kg)要低[26],这可能是由于研究区位于中国北部典型的荒漠生态系统,面临着严峻的水土流失,从而造成了较低的C含量水平[27]。与有机碳、全氮不同,各点的全磷值均高于中国水平(0.56 g/kg)[24],低于全球水平(2.80 g/kg)[23],這说明处于生长中期的胡杨几乎不受P元素的限制。较稳定的P含量和较低的C含量,使得肥岛中的 C/P 值远小于136的中国平均值,且从肥岛外至冠幅下,随着距离的增加C/P值明显升高,表明C/P受到了肥岛效应的显著影响。
土壤C/P和C/N是反映土壤养分发育状况的良好指标。C/P<200意味着养分净矿化,C/P>300意味着净固定,C/P在200~300之间表示可溶性磷含量变化不大,本研究中胡杨冠下肥岛C/P值在3~32,表明土壤微生物体有机磷出现了净矿化现象,土壤磷素的有效性较高。土壤C/N反映了土壤生物分解过程中碳、氮循环与土壤肥力的潜在贡献之间的关系[28],较高的C/N(>25)表明有机质的积累速度快于分解速度。本研究中土壤C/N值处于较低的水平,在10~17,这与Elisabeth等的结果[29]相似,主要是由于C、N这2种元素相对较高的相关系数(0.87)使得C/N受到了较大的约束造成的,表明土壤微生物对有机质的矿化分解速率较快,有机质分解完全,有效氮供应量增加,不利于有机质的积累。而胡杨冠下土壤C/N并未随着距离的改变呈现出明显的变化,表现出一定的稳定性,说明肥岛效应对C/N无显著影响,这可能是由于植物是土壤中C、N 2种元素共同的主要来源,胡杨下土壤C、N的变化较为一致引起的[30]。Koerselman和Meuleman通过总结多种生态系统的试验结果得出结论,当N/P <14,表明是N素限制,N/P>16则是P素限制,N/P处于二者之间为N、P元素共同限制,这个结论被广泛地应用于生态系统养分限制因子的判断[31]。本研究中,N/P最大值为2.58,表明胡杨冠下肥岛存在着较为严重的N素限制,这与上述结论一致。此外,根据生长速率假说(GRH),快速生长的植物生物量N/P通常会很低。对于本研究中处于生长中期的胡杨来说,较快的生长速率导致胡杨对限制性元素N的需求增加,促使胡杨加强对土壤中N的吸收,因此,从肥岛外到冠幅中央N含量的增加速度明显大于P,导致N/P值迅速上升。这表明,随着肥岛N元素富集程度的增强,胡杨冠下土壤N/P发生明显变化,N/P受到肥岛效应的显著影响。
综上所述,在胡杨肥岛的作用下,土壤养分,尤其C元素和限制性N元素向冠幅中央迅速积累,促进了胡杨对周边环境的适应,但也显著改变了胡杨冠下土壤的生态化学计量特征,表明肥岛效应会对土壤生态化学计量产生明显的影响。本试验仅研究单棵胡杨冠下肥岛效应对土壤生态化学计量产生的影响,对于更大尺度上,如群落尺度上这种影响是否存在还不清楚,需要进一步研究。
本研究通过探讨干旱区胡杨肥岛对土壤C、N、P含量及其化学计量特征的影响,得出以下结论:(1)在水平方向和垂直方向上,从肥岛外至冠幅中央随着距离和深度的变化,C、N含量明显增加,表现出明显的肥岛现象,而P变化不明显;(2)N/P和C/P受到了肥岛效应的显著影响,从肥岛外至冠幅中央明显增加,在垂直方向上也受肥岛效应影响随深度增加逐渐减小;(3)胡杨肥岛的P素有效性较高,有机质的矿化分解速率较快,分解完全,且表现出较为严重的N素限制。
参考文献:
[1]Jirk u? V,Kodesova R,Nikodem A,et al. Temporal variability of structure and hydraulic properties of topsoil of three soil types[J]. Geoderma,2013,204(4):43-58.
[2]瞿王龙,杨小鹏,张存涛,等. 干旱、半干旱地区天然草原灌木及其肥岛效应研究进展[J]. 草业学报,2015,24(4):201-207.
[3]Aagren G I. Stoichiometry and nutrition of plant growth in natural communities[J]. Annual Review of Ecology Evolution & Systematics,2008,39(1):153-170.
[4]程 滨,赵永军,张文广,等. 生态化学计量学研究进展[J]. 生态学报,2010,30(6):1628-1637.
[5]喻林华,方 晰,项文化,等. 亚热带4种林分类型枯落物层和土壤层的碳氮磷化学计量特征[J]. 林业科学,2016,52(10):10-21.
[6]Jordi S,Josep P. The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system[J]. Plant Physiology,2012,160(4):1741-1761.
[7]Zhang Z S,Song X L,Lu X G,et al. Ecological stoichiometry of carbon,nitrogen,and phosphorus in estuarine wetland soils:influences of vegetation coverage,plant communities,geomorphology,and seawalls[J]. Journal of Soils and Sediments,2013,13(6):1043-1051.
[8]Wei Z,Liu W C,Xu M P,et al. Response of forest growth to C ∶ N ∶ P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau,China[J]. Geoderma,2019,337(3):280-289.
[9]高丽倩,赵允格,许明祥,等. 生物土壤结皮演替对土壤生态化学计量特征的影响[J]. 生态学报,2018,38(2):678-688.
[10]Xiao L,Peng L,Peng S,et al. Soil nutrient stoichiometries and enzymatic activities along an elevational gradient in the dry-hot valley region of southwestern China[J]. Archives of Agronomy and Soil Science,2019,65(3):322-333.
[11]李 婷,邓 强,袁志友,等. 黄土高原纬度梯度上的植物与土壤碳、氮、磷化学计量学特征[J]. 环境科学,2015,36(8):2988-2996.
[12]Stone M M,Deforest J L,Plante A F. Changes in extracellular enzyme activity and microbial community structure with soil depth at the luquillo critical zone observatory[J]. Soil Biology and Biochemistry,2014,75(4):237-247.
[13]李 磊,李艷红,陈成贺日,等. 1972—2011年期间艾比湖面积变化研究[J]. 湿地科学,2014,12(2):263-267.
[14]崔德宝,于瑞德,于 洋,等. 新疆艾比湖自然保护区阿奇克苏河沿岸胡杨年龄结构和生长量状况研究[J]. 干旱区资源与环境,2012,26(1):194-200.
[15]Agren G I,Hyvnen R,Berglund S L,et al. Estimating the critical N ∶ C from litter decomposition data and its relation to soil organic matter stoichiometry[J]. Soil Biology & Biochemistry,2013,67(6):312-318.
[16]王小平,张 飞,杨胜天,等. 基于三维荧光技术的艾比湖流域地表水盐分快速诊断研究[J]. 光谱学与光谱分析,2018,38(5):1468-1475.
[17]Heba E,Mohamed A,Abu-Alkheir A,et al. Spatial variation of soil carbon and nitrogen pools by using ordinary Kriging method in an area of North Nile Delta,Egypt[J]. Catena,2014,113(3):70-78.
[18]Klemmedson J O,Barth R C. Distribution and balance of biomass and nutrients in desert shrub ecosystems[J]. Reports of Progess,1975:75-85.
[19]Cross A F,Schlesinger W H. Plant regulation of soil nutrient distribution in the northern Chihuahuan Desert[J]. Plant Ecology,1999,145(1):11-25.
[20]刘任涛. 沙地灌丛的“肥岛”和“虫岛”形成过程、特征及其与生态系统演替的关系[J]. 生态学杂志,2014,33(12):3463-3469.
[21]向泽宇,张 莉,张全发,等. 青海不同林分类型土壤养分与微生物功能多样性[J]. 林业科学,2014,50(4):22-31.
[22]Aponte C M T,Season P F. Canopy cover and soil depth[J]. Biogeochemistry,2010,101(5):77-92.
[23]Tian H Q,Chen G S,Zhang C,et al. Pattern and variation of C ∶ N ∶ P ratios in Chinas soils:a synthesis of observational data[J]. Biogeochemistry,2010,98(1/2/3):139-151.
[24]Qing Y S,Li Y. Analysis of soil carbon,nitrogen and phosphorus in degraded alpine wetland,Zoige,southwest China[J]. Acta Prataculturae Sinica,2015,24(2):38-47.
[25]邓玉峰,田善义,成艳红,等. 模拟氮沉降下施石灰对休耕红壤优势植物根际土壤微生物群落的影响[J]. 土壤学报,2019,56(6):1449-1458.
[26]Tao Y,Liu Y B,Wu G L,et al. Regional-scale ecological stoichiometric characteristics and spatial distribution patterns of key elements in surface soils in the Junggar desert,China[J]. Acta Prataculturae Sinica,2016,16(6):5-8.
[27]张彦军,郭胜利,南雅芳,等. 水土流失治理措施对小流域土壤有机碳和全氮的影响[J]. 生态学报,2012,32(18):5777-5785.
[28]Josep P,Sardans J,Rivas-Ubach A,et al. The human-induced imbalance between C,N and P in earths life system[J]. Global Change Biology,2012,18(1):3-6.
[29]Elisabeth N B,Henderson B L. C ∶ N ∶ P stoichiometry in Australian soils with respect to vegetation and environmental factors[J]. Plant and Soil,2013,373(1/2):553-568.
[30]Cleveland C C,Liptzin D. C ∶ N ∶ P stoichiometry in soil:is there a “Redfield ratio” for the microbial biomass?[J]. Biogeochemistry,2007,85(3):235-252.
[31]Koerselman W,Meuleman A F. The vegetation N ∶ P ratio:a new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology,1996,33(6):1441-1450.關健飞,曹 阳. 高通量测序分析黑土稀有微生物群落结构[J]. 江苏农业科学,2020,48(20):288-292.
- 10种杀虫剂防治韭菜迟眼蕈蚊综合药效试验研究
- 南方花椒优质高产综合栽培技术
- 精细化管理在园林施工中的应用
- 黑木耳大棚栽培技术
- 夏秋季利用白叶茶制作卷曲型茶产品工艺研发试验初报
- 浅析茄子开裂原因和防治方法
- 国家玉米新品种展示试验总结报告
- 木醋液对环境的影响及研究进展
- 农业可持续发展中的土壤肥料问题与对策
- 锌肥在水稻节水生产上应用效果对比试验
- 大量元素叶面水溶肥对喀什地区棉花生长及产量 的影响
- 秸秆覆盖技术在果园中的应用
- 玉米春播管理措施
- 水稻栽培技术要点与病虫害防治分析
- 马铃薯春种地膜覆盖高产栽培技术
- 气象服务在农业防灾减灾中的应用
- 关于乡村产业振兴的儿点思考
- 过滤器在节水灌溉中的重要作用
- 浅谈高端智能温室农产品的营销推广
- 以价值评估为导向的河北“田园综合休”发展前景探索
- 俄罗斯和中国在农业领域合作的趋势与前景
- 浅析温室土壤恶化原因及解决办法
- 农业现代化建设中的经济管理问题与对策思考
- 畜禽粪便中残留的抗生素
- 浅谈畜牧养殖中动物疾病防治技术
- elfs
- elicit
- elicitation
- elicitations
- elicited
- eliciting
- elicitor
- elicitors
- elicits
- eligibilities
- eligibility
- eligible
- eligibleness
- eligiblenesses
- eligibles
- eligibly
- eliminabilities
- eliminate
- eliminated
- eliminately
- eliminates
- eliminating
- elimination
- eliminative
- eliminators
- 仁(人)儿苦
- 仂
- 仂语
- 仃
- 仃伶
- 仄
- 仄仄
- 仄仄歪歪
- 仄声
- 仄声韵
- 仄媚
- 仄小
- 仄席
- 仄微
- 仄悚
- 仄慝
- 仄狭
- 仄聞
- 仄行
- 仄道迮径
- 仄陋
- 仄陋狭小
- 仄隘
- 仄韵
- 仅