千里岩岛海域浮游植物种类组成及群落结构特征
孙鹏飞 闫向禄 张晓霞 黄鸿

摘?要:通过2019年11月(秋季)和2020年5月(春季)千里岩岛附近海域两个航次的网采浮游植物调查,共鉴定浮游植物53种,其中硅藻门42种,占79.2%;甲藻门10种,占18.9%;金藻门1种,占1.9%。秋季优势种包含11种,伏氏海线藻(Thalassionema frauenfeldii)为第一优势种;春季优势种包含6种,夜光藻(Noctiluca scientillans)为第一优势种。秋季浮游植物细胞密度平均值为1.27×105 cell/m3,显著高于春季的0.43×104 cell/m3(P<0.01)。千里岩岛附近海域浮游植物细胞丰度较高,这可能是由于陆地营养盐经地表径流输入而促进近岸海域浮游植物的繁殖生长。2019年秋季千里岩岛近岸海域浮游植物的多样性、均匀度和丰富度指数低于离岸海域;2020年春季千里岩岛西北侧调查站位极低的多样性、丰富度和均匀度指数是由于夜光藻的大量繁殖引起的。秋季的多样性和丰富度指数显著高于春季(P<0.01),但均匀度指数在两季间无显著差异(P>0.05)。
关键词:浮游植物;细胞密度;优势种;千里岩岛海域
浮游植物是海洋生态的初级生产者,是许多浮游动物和滤食性贝类的主要饵料,在海洋生态系统物质循环和能量转化过程中具有十分重要的作用[1-2]。自然的或人为的海洋环境变化都会导致浮游植物群落结构变化,故浮游植物群落结构对海洋环境变化具有指示作用[3]。研究浮游植物群落结构对了解海洋生态系统动力过程和渔业资源动态变化具有重要意义[4]。
千里岩岛(北纬36°15′57″,东经121°23′09″)位于南黄海之中,距陆地最近点46 km,岛屿生态系统近二十年几乎无人为干扰,是我国少数几个未被开发的刺参和皱纹盘鲍原种产地之一,同时也是多种黄渤海重要渔业资源的繁育场和索饵场[5]。千里岩岛周围海域为国家级水产种质资源保护区和海洋生态系统省级自然保护区,保持了海洋生态系统的完整性与自然性。目前关于千里岩岛海域浮游生物的研究较少,开展这方面的研究,对该海域生态系统的保护和健康发展具有重要意义。
本研究基于2019年11月(秋季)和2020年5月(春季)在千里岩岛海域网采浮游植物调查数据,分析浮游植物种类组成、细胞密度和群落结构参数时空分布,为该海域生物多样性保护提供基础资料,并为千里岩岛海洋生态系统自然保护区生态系统服务功能的价值评价提供基础数据和参考依据。
1?材料和方法
1.1?研究区域和数据采集
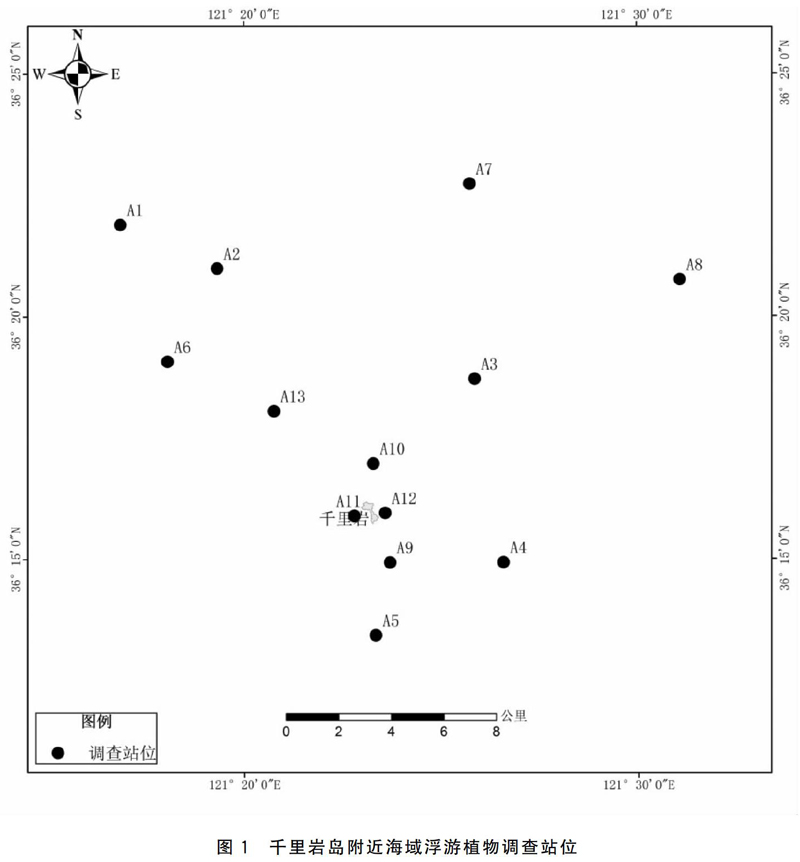
研究区域位于千里岩岛邻近海域(东经121°17′-121°31′,北纬36°13′-36°23′),共设置13个调查站位,千里岩岛北侧设置8个站位,南侧设置5个站位,站位分布如图1所示。A12站位距千里岩岛最近,约0.30 km;A8站位距千里岩岛最远,约15 km。调查时间为2019年11月(秋季)和2020年5月(春季)。浮游植物使用浅水Ⅲ型浮游生物网(网长280 cm、网口内径37 cm,网口面积0.1 m2,网筛绢规格0.077 mm)自底至表做垂直拖网采集,并使用Hydro-bios流量计记录滤水量。浮游植物网采样品均用5%甲醛溶液固定后带回实验室,利用显微镜进行种类的鉴定与计数。样品的采集和分析均按《海洋调查规范》(GB/T 12763-2007)[6]进行。
1.2?分析方法
计算浮游植物群落香农-威纳(Shannon-Weaner)多样性指数(H)、Margalef物种丰富度指数(D)、Pielou均匀度指数(J)值及各物种的优势度(Y),计算公式如下:
式中:S为样品中的种类总数,Pi为第i种个体数与样品总个体數比值,N为样品中的生物个体数,Hmax为log2S,表示多样性指数的最大值。
式中:ni为群落中即所有调查站位第i种的个体总数,N为群落中所有物种的总个体数,fi为第i种个体在各样品中的出现频率。在调查海域以优势度Y>0.02的种类定为优势种[7]。
使用K-W秩和检验方法检验不同季节浮游植物细胞密度、群落结构参数(H、D、J)差异的显著性。数据分析在R软件平台上完成,使用Vegan包计算浮游植物群落结构参数值。
2?结果
2.1?浮游植物种类组成
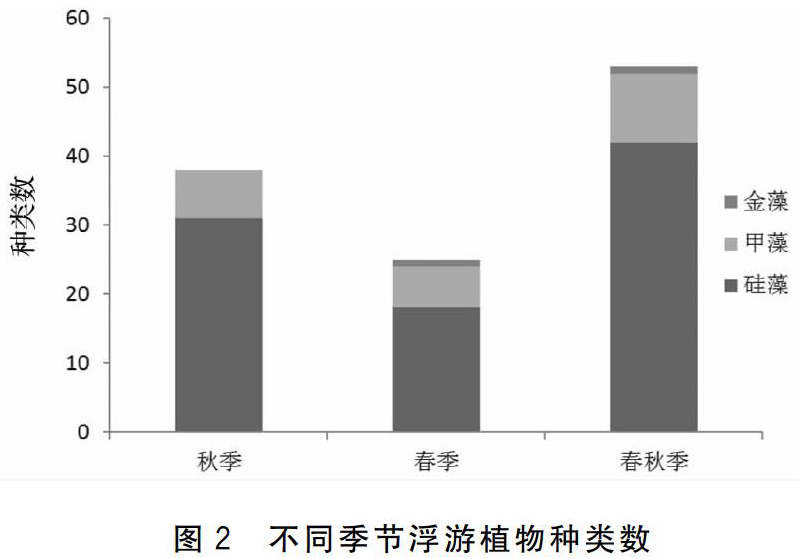
千里岩岛附近海域春秋两季网采的浮游植物共鉴定出2门53种,其中硅藻门42种,占79.2%;甲藻门10种,占18.9%;金藻门1种,占1.9%。2019年秋季鉴定浮游植物38种,2020年春季鉴定浮游植物25种,各季节浮游植物种类组成如图2所示。
2.2?浮游植物细胞密度
2019年秋季浮游植物细胞密度平均值为1.27×105 cell/m3,最低值位于A3站位,为0.65×105 cell/m3;最高值位于A8站位,为2.46×105 cell/m3。A8站位附近海域浮游植物细胞密度较高,千里岩岛正北海域浮游植物细胞密度较低(图3)。2020年春季浮游植物细胞密度平均值为0.43×104 cell/m3,最低值位于S9站位,为0.02×104 cell/m3;最高值位于S11站位,为0.90×104 cell/m3。距千里岩最近的3个调查站位浮游植物丰度较高,千里岩岛南侧海域浮游植物细胞密度较低(图4)。2019年秋季浮游植物细胞密度显著高于2020年春季(P<0.01),高于1个多数量级。
2.3?浮游植物群落结构
2019年秋季浮游植物多样性指数平均值为2.09,A6站位多样性最高,A11站位多样性最低,距千里岩岛较近调查站位的多样性指数较低(图5a);2020年春季浮游植物多样性指数平均值为1.37,显著低于2019年秋季(P<0.01),S9站位多样性最高,S7站位多样性最低,千里岩岛西北侧调查站位的多样性指数较低(图5b)。2019年秋季浮游植物丰富度指数平均值为1.08,A2站位丰富度最高,A5站位丰富度最低,距千里岩岛较近站位的丰富度较低(图6a);2020年春季丰富度指数平均值为0.82,显著低于2019年秋季(P<0.01),距千里岩岛较近站位丰富度较高(图6b)。2019年秋季浮游植物均匀度指数均值为0.81,A7站位均匀度最高,A11站位均匀度最低,千里岩岛较近站位的均匀度较低(图7a);2020年春季均匀度均值为0.68,与2019年春季无显著差异(P>0.05),S4站位均匀度最高,S3站位均匀度最低,千里岩岛西北侧调查站位的均匀度指数较低(图7b)。
2.4?浮游植物群落优势种
2019年秋季浮游植物优势种为圆筛藻、辐射圆筛藻、布氏双尾藻、翼鼻状藻、尖刺伪菱形藻、粗根管藻、伏氏海线藻、叉状角藻、梭形角藻、三角角藻和夜光藻,伏氏海线藻为第一优势种(见表1),细胞丰度均值为2.78×104 cell/m3,优势度为0.135;2020年春季优势种为窄隙角毛藻、辐射圆筛藻、圆筛藻、大角角藻、夜光藻、锥形原多甲藻,夜光藻为第一优势种(见表2),细胞丰度均值为0.13×104 cell/m3,优势度为0.298。3?讨论
浮游植物细胞密度空间分布图显示千里岩岛邻近海域的浮游植物细胞丰度较高,这可能是由于千里岩岛上的营养盐经地表径流输入到海洋中,进而促进千里岩岛近岸海域浮游植物繁殖、生长。张亮等在研究2015年秋季千里岩岛海域浮游植物群落结构特征时,也发现近岸的浮游植物的密度较高,远岸浮游植物密度低[8]。在进行浮游植物调查时,也同步进行了浮游动物调查,调查结果显示2020年春季浮游动物丰度显著高于2019年秋季。浮游动物以浮游植物为食,2020年春季较低的浮游植物丰度可能是由于浮游动物的捕食效果引起的。
浮游植物群落结构评价的三大指数可在一定程度上指示海域环境状况,常以H′<1、J′<0.3作为群落多样性较差的标准进行综合评价[9]。2020年春季H3、H7站位极低的多样性和均匀度指数是由于在这两个站位只鉴定发现5种浮游植物,并且夜光藻(Noctiluca scientillans)细胞丰度显著高于其它种类。夜光藻是海洋赤潮主要发生藻,是冷水种,易斌等在研究2009年至2016年华南近海赤潮分布特征时,发现夜光藻集中在冬季和初春水体温度较低时爆发赤潮[10]。海水温度、硝氮、亚硝氮、COD、磷酸盐等因子对夜光藻生长繁殖都有重要影响[10-12]。尽管本次调查的夜光藻细胞丰度较低,但对千里岩岛海域进行连续的海洋环境监测对海域海洋生态系统的健康发展及发挥自然保护区的服务功能具有重要意义。
参考文献:
[1] 刘长东,郭晓峰,唐衍力,等.海州湾前三岛人工鱼礁区浮游植物群落组成及与环境因子的关系[J].中国水产科学,2015,22(3):545-555.
[2] 喻龙,王磊,王文君,等.庙岛群岛海域网采浮游植物种类组成及分布[J].海洋科學进展,2017,35(3):404-413.
[3] Round F E.The Ecology of Algae[M].London:Cambridge University Press,1981:1-653.
[4] 蔡文贵,李纯厚,贾晓平,等.粤西海域浮游植物群落结构特征的多元分析与评价[J].水生生物学报,2007,3l(2):l55-161.
[5] 宁璇璇,夏炳训,陈权文,等.2013年夏季千里岩岛水产种质资源保护区浮游生物的生态特征[J].海洋通报,2014,33(5):593-598.
[6] 国家质量监督检验检疫局. 海洋调查规范:GB/T 12763.2007[S].北京:中国标准出版社,2007.
[7] 孙儒泳.动物生态学原理[M].2版.北京:北京师范大学出版社,1992.
[8] 张亮,宋春丽,陶卉卉,等.2015年秋季千里岩岛海域浮游植物群落结构特征[J].广西科学院学报,2018,34(2):121-124.
[9] 马建新,郑振虎,李云平,等.莱州湾浮游植物分布特征[J].海洋湖沼通报,2002(4):64-67.
[10] 易斌,陈凯彪,周俊杰,等.2009年至2016年华南近海赤潮分布特征[J].海洋湖沼通报,2018(2):23-31.
[11] 郑向荣,付仲,郗艳娟,等.秦皇岛附近海域夜光藻种群密度变化与环境因子的灰关联分析[J].渔业科学进展,2014,35(2):8-15.
[12] 田达玮,宋书群,陈田田,等.胶州湾夜光藻种群动态及其大量繁殖的生态机制[J].海洋与湖沼,2017,48(2):276-284.
